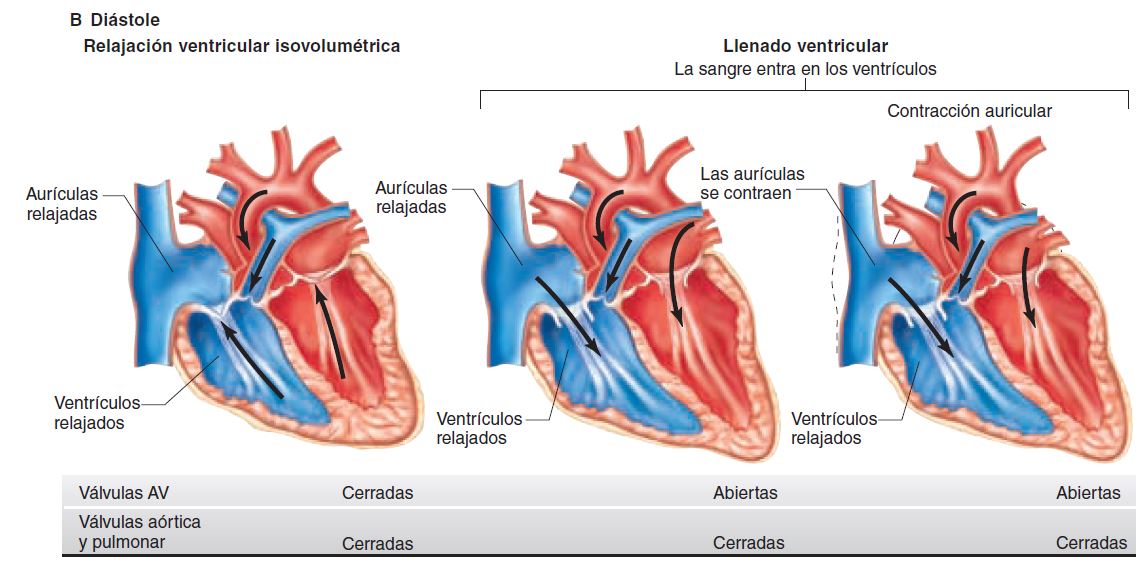
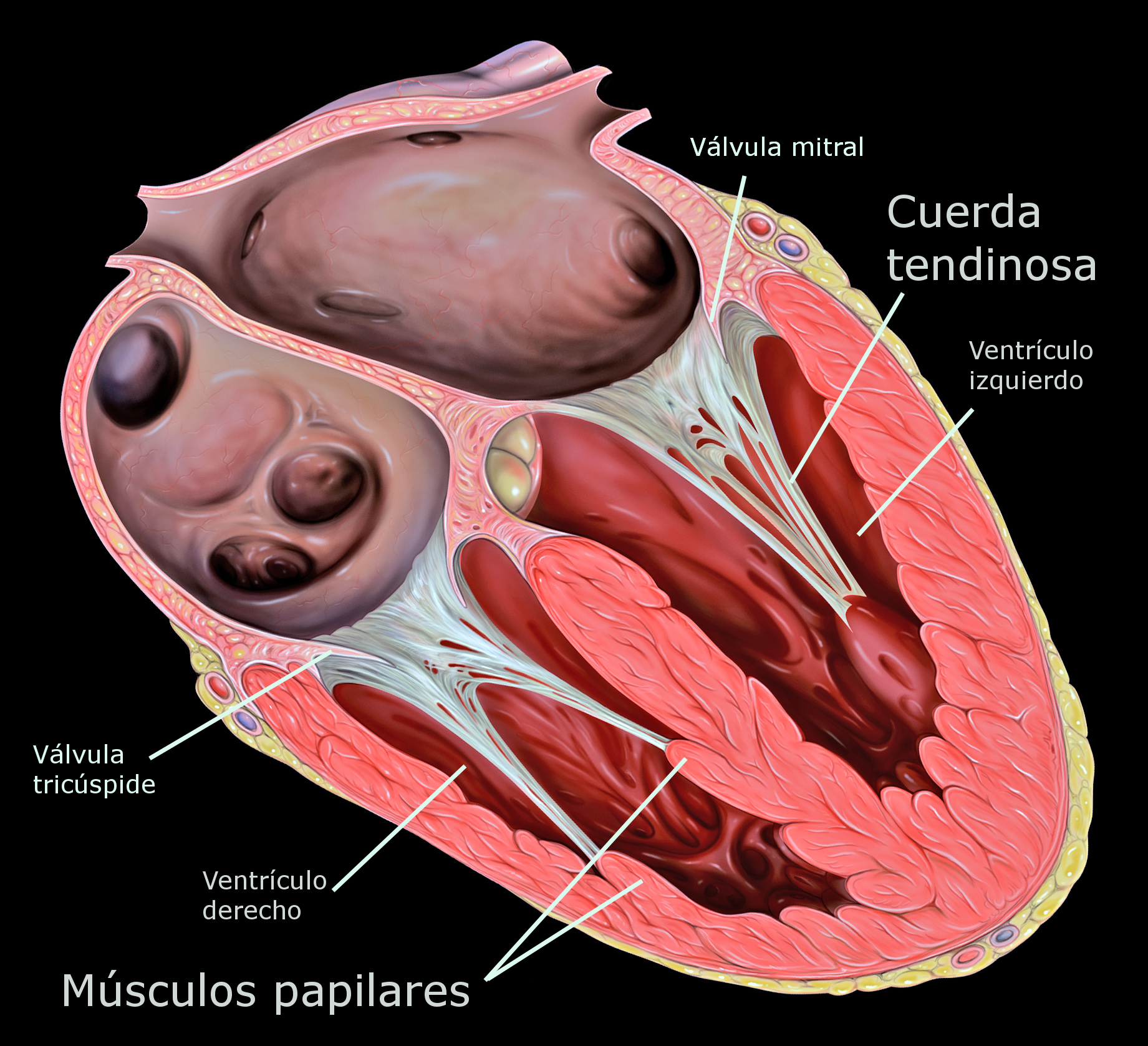
LA MICROCIRCULACIÓN Y EL SISTEMA LINFÁTICO: INTERCAMBIO DE LÍQUIDO CAPILAR, LÍQUIDO INTERSTICIAL Y FLUJO LINFÁTICO.
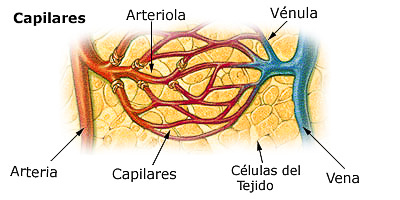
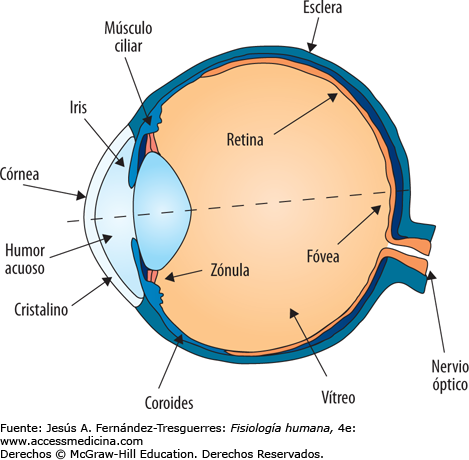
Las arteriolas, que, en general, tienen diámetros internos de solo 10-15m. Entonces las arteriolas se ramifican entre dos y cinco veces, alcanzando diámetros de 5 a 9 m en sus extremos cuando aportan la sangre a los capilares. Las metaarteriolas (las arteriolas termina- les) no tienen una capa muscular continua, sino fibras musculares lisas rodeando el vaso en puntos intermitentes. La pared está compuesta por una capa unicelular de células endoteliales y está rodeada por una membrana basal muy fina en el exterior del capilar. El grosor total de la pared capilar es de solo unas 0,5 m, el diámetro interno del capilar es de 4-9 m, apenas suficiente para el paso de los eritrocitos y otras células sanguíneas exprimidas
Tipos especiales de «poros» en los capilares de algunos órganos
En el cerebro, las uniones entre las células endoteliales capilares son principalmente uniones «estrechas» que permiten la entrada y salida de moléculas muy pequeñas como agua, oxígeno y dióxido de carbono en los tejidos cerebrales.
En el hígado sucede lo contrario. Los espacios entre las células endoteliales capilares son aperturas amplias
Los poros de las membranas capilares gastrointestinales son intermedios entre las de los músculos y las del hígado.
El flujo sanguíneo a través de cada capilar es intermitente, hay tantos capilares en los tejidos que su función global termina por ser superada, es decir, hay una velocidad media del flujo sanguíneo a través de cada lecho capilar tisular, una presión capilar media dentro de los capilares y una velocidad de transferencia media de las sustancias entre la sangre de los capilares y el líquido intersticial circundante.
Difusión a través de la membrana capilar
Con mucho, el medio más importante por el cual se transfieren las sustancias entre el plasma y el líquido intersticial es la difusión. Las sustancias hidrosolubles y no liposolubles difunden sólo a través de los «poros» intercelulares en la membrana capilar.
Pero no pueden pasar a través de las membranas lipídicas de las células endoteliales; estas sustancias son las propias moléculas de agua, los iones sodio y cloruro y la glucosa.
la velocidad con la que difunden las moléculas de agua a través de la membrana capilar es unas 80 veces mayor que la velocidad con la que el propio plasma fluye linealmente
Efecto del tamaño molecular sobre el paso a través de los poros.
La profundidad de los espacios intercelulares capilares, 6 a 7nm, es unas 20 veces el diámetro de la molécula de agua, que es la molécula más pequeña que normalmente atraviesa los poros de los capilares. Por el contrario, los diámetros de las moléculas proteicas plasmáticas son ligeramente mayores que la anchura de los poros.
Intersticio y líquido intersticial
Una sexta parte del volumen total del organismo consiste en espacios entre las células, que colectivamente se conoce como el intersticio. El líquido de estos espacios es el líquido intersticial.
Gel en el intersticio
El líquido del intersticio deriva por filtración y difusión de los capilares. Esta combinación de filamentos de proteoglicano y líquido atrapado dentro de ellos tiene las características de un gel y, por tanto, se conoce como gel tisular. Debido al gran número de filamentos de proteoglicano, es difícil que el líquido fluya fácilmente a través de este gel tisular.
Líquido «libre» en el intersticio.
Aunque casi todo el líquido del intersticio está atrapado dentro del gel tisular, a veces también hay pequeños riachuelos de líquido «libre» y pequeñas vesículas de líquido libre, lo que significa que carece de moléculas de proteoglicano y, por tanto, puede fluir libremente.
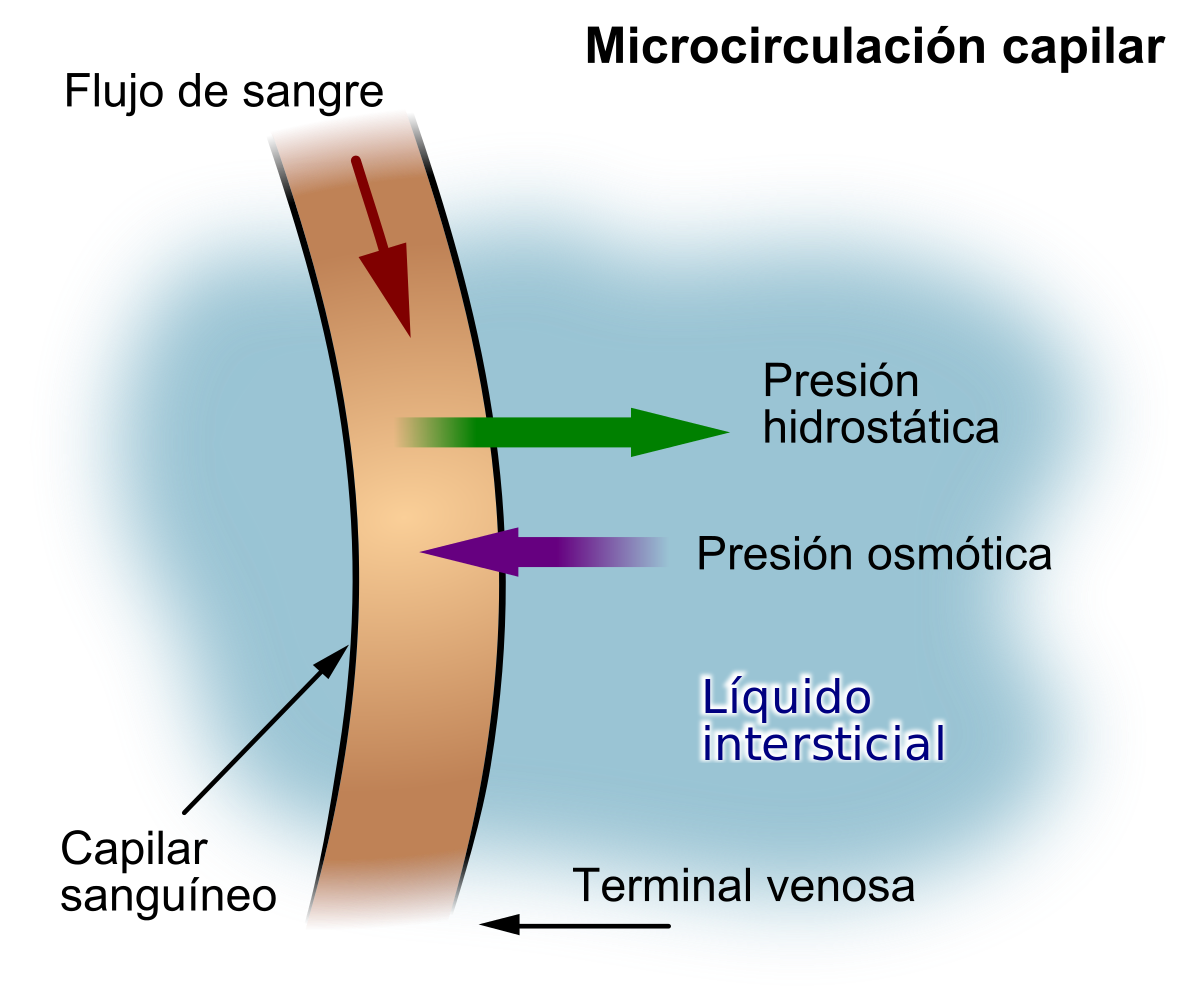
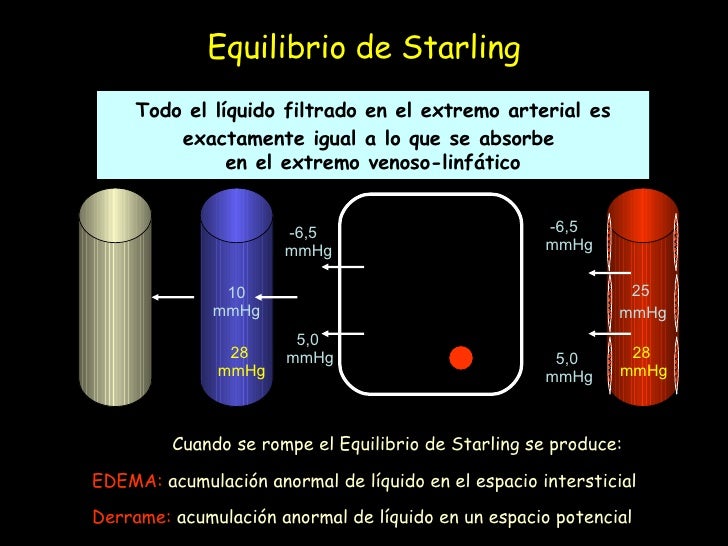
Equilibrio de Starling
Para el intercambio capilar la cantidad de líquido que se filtra de los extremos arteriales de los capilares hacia el exterior es casi exactamente igual a la de líquido que vuelve a la circulación mediante absorción. Se obtiene la media de las presiones de los capilares arteriales y venosos para calcular la media de la presión capilar funcional a lo largo de todo el capilar. Se calcula que es de 17,3 mmHg. n cuanto a la circulación capilar total encontramos un equilibrio casi perfecto entre las fuerzas totales de salida, 28,3 mmHg, y la fuerza total de entrada, 28,0 mmHg. Este ligero desequilibrio de fuerzas, 0,3 mmHg
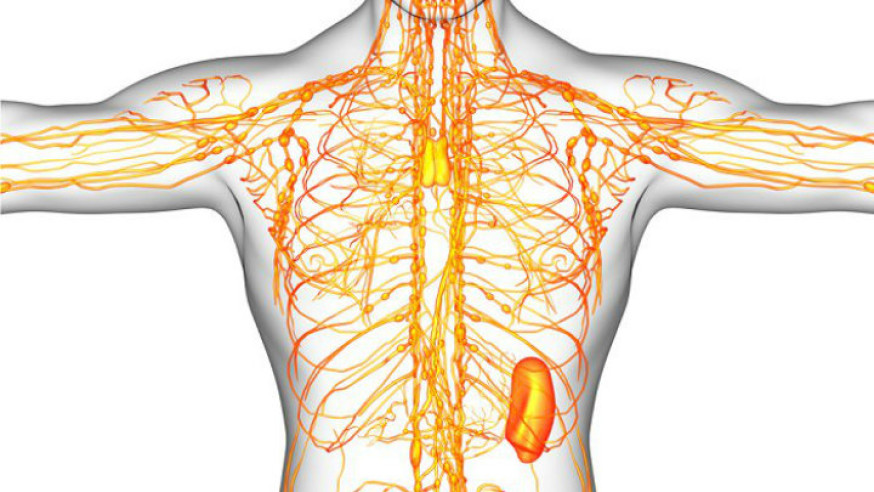
Sistema linfático
Una vía accesoria a través de la cual el líquido puede fluir desde los espacios intersticiales hacia la sangre. Es más, los linfáticos transportan las proteínas y las macropartículas de los espacios tisulares, ya que ninguna de las cuales podrá ser eliminada por absorción directamente hacia los capilares sanguíneos
Capilares linfáticos terminales y su permeabilidad.
La mayoría del líquido que se filtra desde los extremos arteriales de los capilares sanguíneos fluye entre las células y, por último, se reabsorbe de nuevo hacia los extremos venosos de los capilares sanguíneos. El líquido que vuelve a la circulación a través de los linfáticos es muy importante porque las sustancias de alto peso molecular que contiene, como las proteínas, no pueden ser absorbidas desde los tejidos de ninguna otra forma. En ella se muestran las células endoteliales de los capilares linfáticos unidos mediante filamentos que se anclan en el tejido conjuntivo circundante.
los linfáticos contienen válvulas en los extremos de los capilares linfáticos terminales y también en el recorrido de los vasos mayores hasta el punto en que se vacían en la circulación sanguínea.
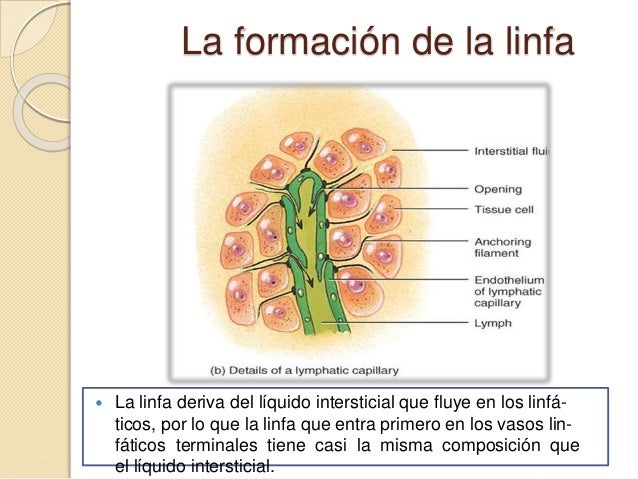
Formación de la linfa
La linfa deriva del líquido intersticial que fluye en los linfáticos, por lo que la linfa que entra primero en los vasos linfáticos terminales tiene casi la misma composición que el líquido intersticial.
Velocidad del flujo linfático
En un ser humano en reposo pasan 100 ml por hora en el flujo linfático a través del conducto torácico, y otros 20 ml fluyen hacia la circulación cada hora a través de otros canales, con un total del flujo linfático estimado en torno a 120 ml/h o 2-3 l al día.
¿Qué funciones tienen lugar en la microcirculación?
El transporte de nutrientes a los tejidos
La eliminación de los residuos celulares.
Las arterias pequeñas controlan el flujo sanguíneo en cada área tisular y las condiciones locales en los tejidos controlan los diámetros de las arterias.
Todas las anteriores
Ninguna de las anteriores.
¿Qué son los capilares y que se produce en ellos?
Los capilares son estructuras extremadamente finas
Tienen paredes de varias capas de células endoteliales poco permeables
Produce el intercambio de nutrientes y residuos celulares entre los tejidos y la sangre circulante.
Solo a y c son correctas
Ninguna es correcta.
¿Cómo es la estructura de la microcirculación y de sistema capilar?
Es específica para servir a sus necesidades especiales
Es la misma para todos los órganos
Solo se ramifica de 4-6 veces
El diámetro de las arteriolas es de 25 micrómetros
Todas son correctas
¿Qué estructura de la microcirculación tiene músculos y cómo se distribuyen los mismos?
Arteriolas
Metaarteriolas
Vénulas
Todas
Ninguna
¿A qué se le denomina vasomotilidad?
Al fluido continuo de sangre a los capilares
Al flujo intermitente cada minuto o segundo de la sangre a los capilares
Ambos
Ninguna
¿Cómo es la regulación de la vasomotilidad?
Si el consumo de oxígeno es alto, los periodos intermitentes de flujo sanguíneo son frecuentes y prolongados.
Si el consumo de oxígeno es alto, no hay periodos intermitentes
Si el consumo de oxígeno es alto solo se vuelve más prolongado
A y c son correctas
Ninguna es correcta.
¿cómo es la difusión a través de la membrana capilar?
Difusión
Osmosis
Ambas
Ninguna.
Características especiales de los capilares linfáticos.
Parte del líquido filtrado se reabsorbe en los extremos venosos de los capilares sanguíneos
Una décima parte del líquido entra en los capilares sanguíneos
La cantidad de linfa total es de 5-5 litros por día
A y b son correctas
Ninguna es correcta.
¿cuáles son los factores básicos que determinan el movimiento del líquido a través de la membrana capilar?
Presión capilar
Presión del líquido intersticial
Presión coloidosmótica del plasma
Presión coloidosmótica del líquido intersticial
Todas son correctas.
¿qué factores aumentan la presión intersticial?
Presión capilar elevada
Baja permeabilidad de los capilares
Presión coloidosmótica del plasma alta
A y c son correctas
Todas sin correctas
¿Qué factores externos originan el bombeo linfático?
Contracción de los músculos vecinos
Movimiento de partes del cuerpo
Pulsaciones arteriales
Compresión de los tejidos por objetos externos
Todas son correctas
¿Cuáles son los 2 factores primarios que determinan el flujo de linfa?
La presión del plasma
La presión del liquido intersticial
La actividad de la bomba linfática
B y c son correctas
A y b son correctas
¿Qué significa que una sustancia sea liposoluble?
Es de fácil digestión
Que se difunde por medio de quilomicrones
Que se difunde de manera directa por las paredes celulares
Todas son correctas
Ninguna es correcta
Explica la estructura del intersticio.
Tiene haces de fibra de colágeno
Tiene filamentos de proteoglucano
Ambas son correctas
Ninguna es correcta
¿qué efecto tiene la presión en los capilares?
Ninguna
Fuerza el líquido fuera de ellos
Fuerza el líquido por los poros capilares
B y c son correctas
Valores normales para la presión coloidosmótica del plasma.
28-19mmHG
10-22mmHG
12-15mmHG
5-10mmHG
27-30mmHG
¿Cuál es la concentración media de proteínas en el líquido intersticial?
50%
45%
40%
42.5%
40.6%
¿Dónde drena la linfa de la región inferior del cuerpo?
Drena en la VCI
Drena en el conducto linfático derecho
Drena en la unión de la vena yugular interna izquierda y la vena subclavia
Ninguna es correcta
¿Dónde drena la linfa de la región superior del cuerpo?
En el conducto linfático derecho
En la unión de la subclavia derecha y la yugular interna
A y b son correctas
Ninguna es correcta
¿Qué mecanismo falla para que se provoque un edema?
Cuando los tejidos pierden su presión negativa, el líquido se acumula en los espacios y se produce el edema.
Las arteriolas, que, en general, tienen diámetros internos de solo 10-15m. Entonces las arteriolas se ramifican entre dos y cinco veces, alcanzando diámetros de 5 a 9 m en sus extremos cuando aportan la sangre a los capilares. Las metaarteriolas (las arteriolas termina- les) no tienen una capa muscular continua,sino fibras musculares lisas rodeando el vaso en puntos intermitentes.
La pared está compuesta por una capa unicelular de células endoteliales y está rodeada por una membrana basal muy fina en el exterior del capilar. El grosor total dela pared capilar es de solo unas 0,5 m, el diametro interno del capilar es de 4-9 m,apenas suficiente para el paso de los eritrocitos y otras células sanguíneas exprimidas
Tipos especiales de «poros» en los capilares de algunos órganos
1.-En el cerebro, las uniones entre las células endoteliales capilares son principalmente uniones «estrechas» que permiten la entrada y salida de moléculas muy pequeñas como agua, oxígeno y dióxido de carbono en los tejidos cerebrales. 2. En el hígado sucede lo contrario. Los espacios entre las células endoteliales capilares son aperturas amplia 3. Los poros de las membranas capilares gastrointestina- les son intermedios entre las de los músculos y las del hígado.
El flujo sanguíneo a traves de cada capilar es intermitente, hay tantos capilares en los tejidos que su función global termina por ser superada, es decir, hay una velocidad media del flujo sanguíneo a traves de cada lecho capilar tisular, una presión capilar media dentro de los capilares y una velocidad de transferencia media de las sustancias entre la sangre de los capilares y el líquido intersticial circundante.
Difusión a través de la membrana capilar Con mucho, el medio más importante por el cual se transfieren las sustancias entre el plasma y el líquido intersticial es la difusión
Las sustancias hidrosolubles y no liposolubles difunden sólo a través de los «poros»intercelulares en la membrana capilar. Pero no pueden pasar a través de las membranas lipídicas de las células endoteliales; estas sustancias son las propias moléculas de agua, los iones sodio y cloruro y laglucosa. la velocidad con la que difunden las moléculas de agua a través de la membrana capilar es unas 80 veces mayor que la velocidad con la que el propio plasma fluye linealmente
Efecto del tamaño molecular sobre el paso a través de los poros. La profundidad de los espacios intercelulares capilares, 6 a 7nm, es unas 20 veces el diámetro de la molécula de agua, que es la molécula más pequeña que normalmente atraviesa los poros de los capilares. Por el contrario, los diámetros de las moléculas proteicas plasmáticas son ligeramente mayores que la anchura de los poros.
intersticio y líquido intersticial Una sexta parte del volumen total del organismo consiste en espacios entre las células,que colectivamente se conoce como el intersticio. El líquido de estos espacios es el líquido intersticial.
Gel en el intersticio El líquido del intersticio deriva por filtración y difusión de los capilares. Esta combinación de filamentos de proteoglicano y líquido atrapado dentro de ellos tiene las características de un gel y, por tanto, se conoce como gel tisular. Debido al gran número de filamentos de proteoglicano, es difícil que el líquido fluya fácilmente a través de este gel tisular.
Líquido «libre» en el intersticio. Aunque casi todo el líquido del intersticio está atrapado dentro del gel tisular, a veces también hay pequeños riachuelos de líquido «libre» y pequeñas vesículas de líquido libre, lo que significa que carece de moléculas de proteoglicano y, por tanto, puede fluir libremente
Equilibrio de Starling Para el intercambio capilar la cantidad de líquido que se filtra de los extremos arteriales de los capilares hacia el exterior es casi exactamente igual a la de líquido que vuelve a la circulación mediante absorción. Se obtiene la media de las presiones de los capilares arteriales y venosos para calcular la media de la presión capilar funcional a lo largo de todo el capilar. Se calcula que es de 17,3 mmHg. n cuanto a la circulación capilar total encontramos un equilibrio casi perfecto entre las fuerzas totales de salida, 28,3 mmHg, y la fuerza total de entrada, 28,0 mmHg. Esteligero desequilibrio de fuerzas, 0,3 mmHg
Sistema linfático Una vía accesoria a través de la cual el líquido puede fluir desde los espacios intersticiales hacia la sangre. Es más, los linfáticos transportan las proteínas y las macropartículas de los espacios tisulares, ya que ninguna de las cuales podrá ser eliminada por absorción directamente hacia los capilares sanguíneos
Capilares linfáticos terminales y su permeabilidad. La mayoría del líquido que se filtra desde los extremos arteriales de los capilares sanguíneos fluye entre las células y, por último, se reabsorbe de nuevo hacia los extremos venosos de los capilares sanguíneos. El líquido que vuelve a la circulación a través de los linfáticos es muy importante porque las sustancias de alto peso molecular que contiene, como las proteínas, no pueden ser absorbidas desde los tejidos de ninguna otra forma. En ella se muestran las células endoteliales de los capilares linfáticos unidos mediante filamentos que se anclan en el tejido conjuntivo circundante. los linfáticos contienen válvulas en los extremos de los capilares linfáticos terminales y también en el recorrido de los vasos mayores hasta el punto en que se vacían en la circulación sanguínea.
Formacion de la linfa La linfa deriva del líquido intersticial que fluye en los linfáticos, por lo que la linfa que entra primero en los vasos linfáticos terminales tiene casi la misma composición que el líquido intersticial
Velocidad del flujo linfático En un ser humano en reposo pasan 100 ml por hora en el flujo linfático a través del conducto torácico, y otros 20 ml fluyen hacia la circulación cada hora a través de otros canales, con un total del flujo linfático estimado en torno a 120 ml/h o 2-3 l al día.
La función de la circulación consiste en atender las necesidades del organismo: transportar nutrientes hacia los tejidos del organismo, transportar los productos de desecho, transportar las hormonas de una parte del organismo a otra y, en general, mantener un entorno apropiado en todos los líquidos tisulares del organismo para lograr la supervivencia y funcionalidad óptima de las células. Está divida en circulación sistémica y circulación pulmonar:
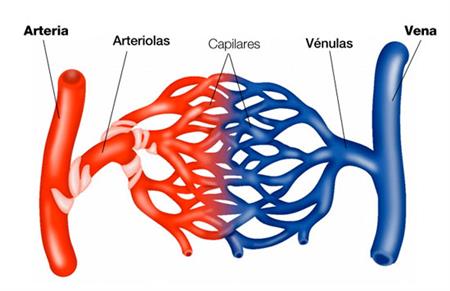
La función de las arterias consiste en transportar la sangre con una presión alta hacia los tejidos, motivo por el cual las arterias tienen unas paredes vasculares fuertes y unos flujos sanguíneos importantes con una velocidad alta. Las arteriolas son las últimas ramas pequeñas del sistema arterial y actúan controlando los conductos a través de los cuales se libera la sangre en los capilares.
Tienen paredes musculares fuertes que pueden cerrarlas por completo o que pueden, al relajarse, dilatarlos vasos varias veces.Los capilares consiste en el intercambio de líquidos, nutrientes, electrólitos, hormonas y otras sustancias en la sangre y en el líquido intersticial. Las paredes del capilar son muy finas y tienen muchos poros capilares diminutos, que son permeables al agua y a otrasmoléculas pequeñas.
Las vénulas recogen la sangre de los capilares y después se reúnen gradualmente formando venas de tamaño progresivamente mayor.
Las venas funcionan como conductos para el transporte de sangre que vuelve desde las vénulas al corazón; igualmente importante es que sirven como una reserva importante desangre extra. Paredes finas.
Volúmenes de sangre en los distintos componentes de la circulación.
Superficies transversales y velocidades del flujo sanguíneo:
Volúmenes de sangre en los distintos componentes de la circulación. Superficies transversales y velocidades del flujo sanguíneo Como debe pasar el mismo volumen de flujo sanguíneo (F) a través de cada segmento dela circulación en cada minuto, la velocidad del flujo sanguíneo (v) es inversamente proporcional a la superficie transversal vascular (A).
v=F/A
En condiciones de reposo la velocidad es como media de 33cm/s en la aorta pero con una velocidad sólo de 1/1.000 en los capilares, es decir, aproximadamente 0,3 mm/s. No obstante, como los capilares tienen una longitud de sólo 0,3 a l mm, la sangre sólo se queda allí durante 1-3s.
Presiones en las distintas porciones de la circulación.
La presión arterial alterna entre una presión sistólica de 120 mmHg y una diastólica de 80mmHg, La presión de los capilares sistémicos oscila desde 35 mmHg cerca de los extremos arteriolar es hasta tan sólo 10 mmHg cerca de los extremos venosos, pero la presión media «funcional» en la mayoría de los lechos vasculares es de 17 mmHg, aproximadamente, una presión suficientemente baja que permite pequeñas fugas de plasma a través de los poros diminutos de las paredes capilares, aunque los nutrientes pueden difundir fácilmente a través de los mismos poros hacia las células de los tejido sexternos. La presión sistólica arterial pulmonar alcanza un promedio de 25 mmHg y la diastólica, de 8 mmHg, con una presión arterial pulmonar media de sólo 16 mmHg. La media de la presión capilar pulmonar alcanza un promedio de sólo 7 mmHg. Aun así, el flujo sanguíneo por minuto a través de los pulmones es el mismo que en la circulación sistémica.
Principios básicos de la función circulatoria
1. La velocidad del flujo sanguíneo en cada tejido del organismo casi siempre se controla por precisión en relación con las necesidades del tejido: Hormonas y sistema nervioso colaboran en el control del flujo sanguíneo tisular. 2. El gasto cardíaco se controla principalmente por la suma de todos los flujos tisulares locales. 3. La regulación de la presión arterial es generalmente independiente del control del flujo sanguíneo local o del control del gasto cardíaco.
Interrelaciones entre la presión, el flujo y la resistencia.
El flujo sanguíneo que atraviesa un vaso sanguíneo está determinado por dos factores: 1)diferencia de presión de la sangre entre los dos extremos de un vaso, también denominado «gradiente de presión» en el vaso, que es la fuerza que empuja la sangre a través del vaso 2) los impedimentos que el flujo sanguíneo encuentra en el vaso,que se conoce como resistencia vascular.
El flujo atraves del vaso se puede calcular con la ley de Ohm que es: ° F=∆P/R ° F= Flujo sanguíneo ° ∆P= Diferencia de presiones de los dos extremos del vaso ° R= resistencia
Flujo sanguíneo
Es la cantidad de sangres que atraviesa un punto dado de la circulación en un periodo de tiempo determinado. ml/m o ml/s.
El flujo sanguíneo global de toda la circulación de un adulto en reposo es de unos 5.000ml/min se considera igual al gasto cardiaco porque es la cantidad de sangre que bombea el corazón en la aorta en cada minuto.
Presión sanguínea
Se mide casi siempre en milímetros de mercurio (mmHg) porque el manómetro de mercurio se ha usado como patrón de referencia para medir la presión
La presión arterial mide la fuerza ejercida por la sangre contra una unidad de superficie dela pared del vaso.En ocasiones, la presión se mide en centímetros de agua (cm H20).
Una presión de 10cm H20 significa una presión suficiente para elevar una columna de agua contra la gravedad hasta una altura de 10 centímetros.
Una presión de 1 mm de mercurio es igual a una presión de 1,36 cm de agua.
Distensibilidad vascular
Todos los vasos sanguíneos del aparato vascular son distensibles, esto permite el acoplamiento de las arterias al gasto pulsátil del corazón y superar las pulsaciones de la presión.
Los vasos más distensibles del cuerpo son las venas, capaces de almacenar 0.5-1 litro de sangre. Las arterias son más fuertes que las venas, es por eso que estas últimas son más distensibles.
La estimulación simpática aumenta la presión en cada volumen de arterias o venas, mientras que la inhibición simpática lo disminuye.
Compliancia diferida: se refiere a que si un vaso está expuesto a un aumento de volumen, primero presentara un incremento de la presión pero progresivamente se estirara el musculo liso del vaso y hará que la presión vuelva a la normalidad.
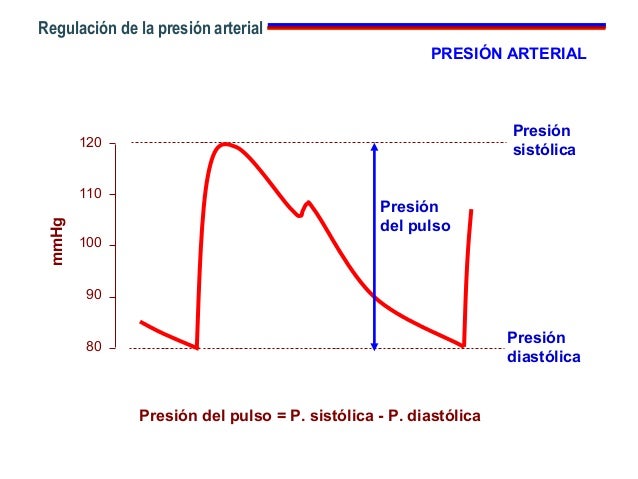
Pulsaciones de la presión arterial
La compliancia del árbol arterial permite reducir las pulsaciones de la presión hasta que prácticamente desaparecen al momento de alcanzar los capilares.
Presión sistólica -> 120 mmHg presión diastólica -> 80 mmHg la diferencia entre estas dos presiones recibe el nombre de presión de pulso y es igual a -> 40mmHg.
Factores que afectan la presión de pulso: 1.- Volumen sistólico del corazón. 2.- Compliancia (distensibilidad total) del árbol arterial (en los ancianos la distensibilidad de los vasos disminuye por la arterioesclerosis). 3.- Eyección del corazón durante la sístole.
La presión de pulso está determinada por la relación entre el gasto cardíaco y la compliancia del árbol arterial.
1.- Estenosis valvular aórtica: se debe a la disminución del diámetro de apertura de la válvula lo cual la presión de pulso aórtico disminuye también.Situaciones que provocan un perfil anormal de la onda de pulso de presión:
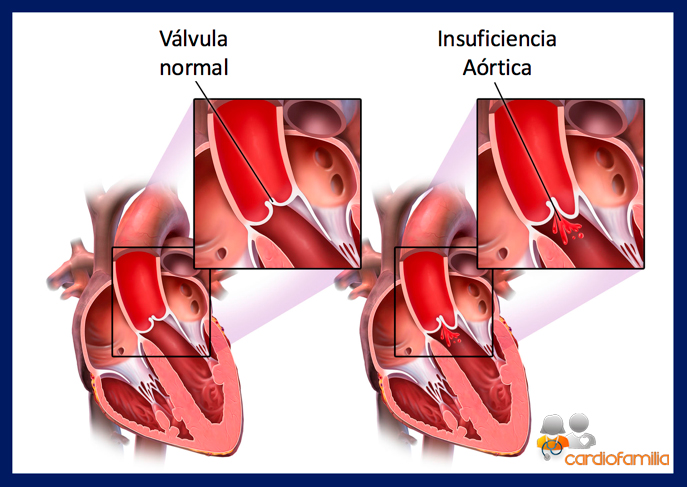
2.- Conducto arterioso permeable: es cuando la sangre en lugar de bombearse a la aorta fluye a través de la arteria y vasos pulmonares, lo que produce un descenso diastólica antes del latido. 3.- Insuficiencia aórtica: es por la ausencia o por el cierre incompleto de la válvula, lo que produce que después de cada latido la sangre que fluye por la aorta se va hacia el ventrículo izquierdo, lo que provoca produce una disminución de la presión aortica.
La velocidad de la transmisión del pulso de la presión en la aorta normal es de 3-5 m/s, de 7-10 m/s en las ramas arteriales grandes y de 15-35 m/s en las pequeñas arterias.
La disminución de las pulsaciones en la periferia recibe el nombre de amortiguación de los pulsos, sus orígenes son: 1.- Resistencia al movimiento de la sangre en los vasos. 2.-Compliancia de los vasos.
El grado de amortiguación es casi directamente proporcional al producto resistencia por compliancia. Se utiliza la arteria antecubital en el método de auscultación para determinar las presiones arteriales. Los ruidos que se escucha reciben el nombre de ruidos de Korotkoff.
Las venas y sus funciones
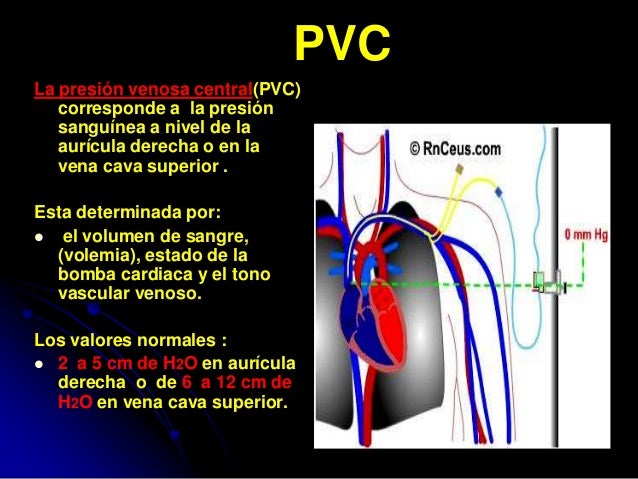
Las venas periféricas pueden impulsar la sangre mediante la bomba venosa e incluso pueden regular el gasto cardíaco. La sangre de todas las venas sistémicas fluyen hacia la aurícula izquierda del corazón. La presión del interior de esta cámara recibe el nombre de presión venosa central.
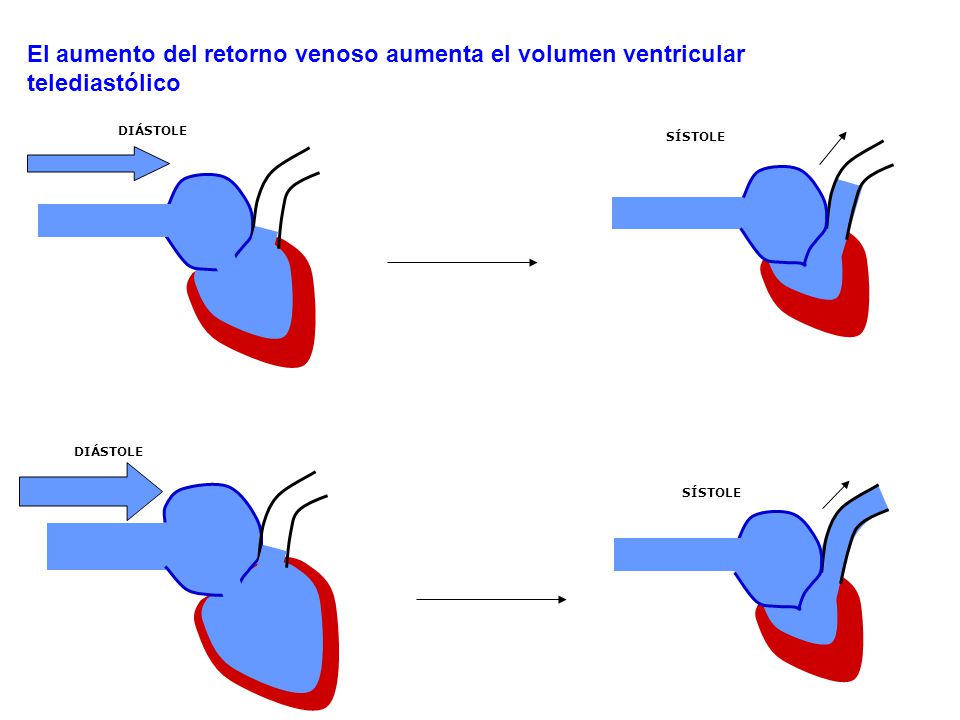
La presión de la aurícula izquierda está regulada por: 1.-La capacidad del corazón de bombear la sangre al exterior de la aurícula y el ventrículo derecho a los pulmones. 2.-La tendencia de la sangre a fluir desde las venas periféricas hacia la aurícula derecha.
Factores que aumentan el retorno venoso (también la presión de la aurícula derecha): 1.-Aumento del volumen de sangre. 2.-Aumento del tono de los grandes vasos del organismo. 3.-Dilatación de las arteriolas.
La presión normal de la aurícula derecha es de 0 mmHg.
Cuando la presión intraabdominal aumenta, la presión de las venas de las piernas debe de aumentar por encima de la presión abdominal antes de que las venas abdominales se abran y permitan el paso de la sangre desde las piernas al corazón.
Presión gravitacional o hidrostática: es producida en el aparato vascular por el peso de la sangre en las venas.
Las venas del cuello de una persona que esté de pie se colapsan casi por completo en todo su recorrido hasta el cráneo, por la presión atmosférica que hay fuera del cuello.
Las venas del interior del cráneo se encuentran dentro de una cámara no colapsable (la cavidad craneal), por lo que no se pueden colapsar. En consecuencia, puede haber una presión negativa en los senos de la dura de la cabeza; en bipedestación la presión venosa del seno sagital de la parte superior del cráneo es de –10 mmHg, por la “aspiración” hidrostática que existe entre la parte superior y la base del cráneo.
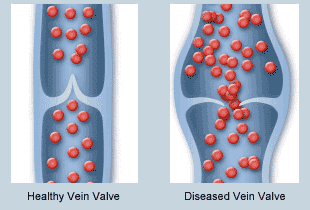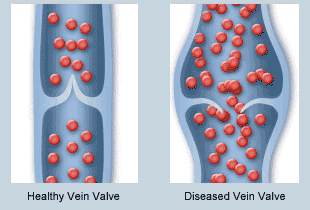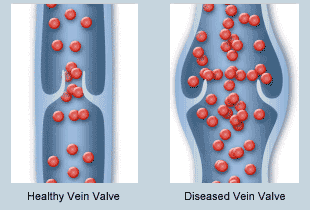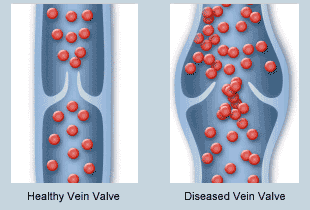
Las válvulas de las venas están distribuidas a tal manera que la dirección del flujo sanguíneo venoso solo puede ir hacia el corazón.
Bomba venosa o bomba muscular: es cuando la tensión de los músculos empuja una determinada cantidad de sangre venosa al corazón.
La presión venosa puede determinarse observando el grado de distensión de las venas periféricas, especialmente la del cuello, o de una manera exacta a través de un catéter a través de las venas periféricas hasta la aurícula derecha.
La presión medida de la aurícula derecha es de 0mmHg y la presión arterial es de 100mmHg.
En la válvula tricúspide o cerca de ella no afectan los factores de presión gravitacional.
El corazón actúa como un regulador de retroalimentación de presión en la válvula tricúspide.
Más del 60% de toda la sangre venosa se encuentra en las venas, es por eso que se dice que actúan como reservorio de energía.
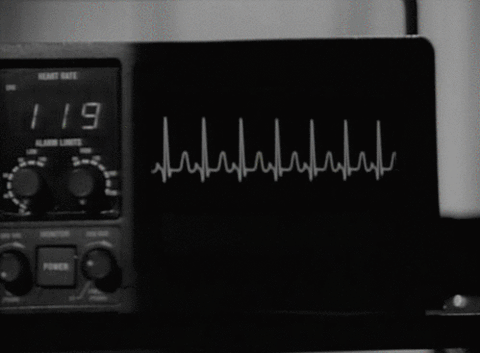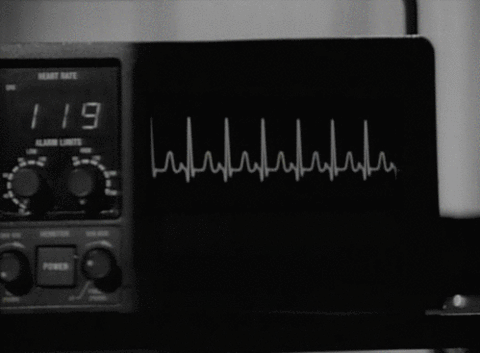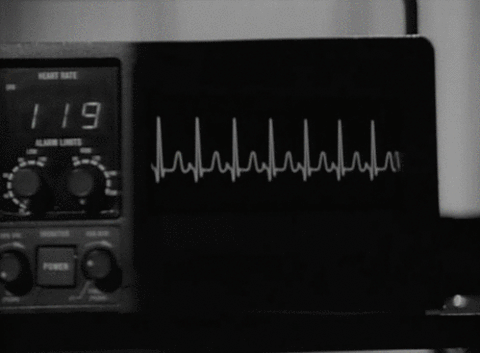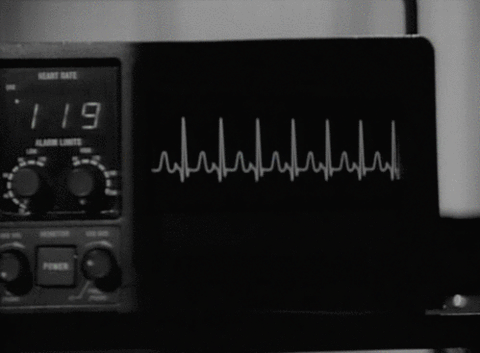
Cuando el impulso cardíaco atraviesa el corazón, la corriente eléctrica también se propaga desde el corazón hacia los tejidos adyacentes que lo rodean. Una pequeña parte de la corriente se propaga hacia la superficie corporal. Si se colocan electrodos en la piel en lados opuestos del corazón se pueden registrar los potenciales eléctricos que se generan por la corriente; el registro se conoce como electrocardiograma (ECG).
CARACTERÍSTICAS DEL ELECTROCARDIOGRAMA NORMAL
El ECG normal está formado por una onda P, un complejo QRS y una onda T. Con frecuencia, aunque no siempre, el complejo QRS está formado por tres ondas separadas: la onda Q, la onda R y la onda S.
La onda P está producida por los potenciales eléctricos que se generan cuando se despolarizan las aurículas antes del comienzo de la contracción auricular. El complejo QRS está formado por los potenciales que se generan cuando se despolarizan los ventrículos antes de su contracción, es decir, a medida que la onda de despolarización se propaga por los ventrículos. Por tanto, tanto la onda P como los componentes del complejo QRS son las ondas de despolarización.
La onda T está producida por los potenciales que se generan cuando los ventrículos se recuperan del estado de despolarización. Este proceso normalmente aparece en el músculo ventricular entre 0,25 y 0,35 s después de la despolarización. La onda T se conoce como onda de repolarización.
Así, el ECG está formado por ondas tanto de despolarización como de repolarización. Los principios de la despolarización y de la repolarización se analizan en el capítulo 5. La distinción entre ondas de despolarización y ondas de repolarización es tan importante en electrocardiografía que requiere una aclaración adicional.
Durante la despolarización el potencial negativo normal del interior de la fibra se invierte y se hace ligeramente positivo en el interior y negativo en el exterior.
Relación del potencial de acción monofásico del músculo ventricular con las ondas QRS y T del electrocardiograma estándar
El potencial de acción monofásico del músculo ventricular, que se ha analizado en el capítulo 10, normalmente dura entre 0,25 y 0,35 s. El potencial de acción monofásico registrado con un microelectrodo insertado en el interior de una fibra muscular ventricular única. El ascenso de este potencial de acción está producido por la despolarización, y la vuelta del potencial al nivel basal está producida por la repolarización.
En un registro simultáneo del ECG de un ventrículo. Las ondas QRS aparecen al principio del potencial de acción monofásico y la onda T aparece al final. Obsérvese específicamente que no se registra ningún potencial en el ECG cuando el músculo ventricular está completamente polarizado o completamente despolarizado. Solo cuando el músculo está parcialmente polarizado o parcialmente despolarizado hay flujo de corriente desde una parte de los ventrículos hacia la otra, y por tanto la corriente también fluye hacia la superficie del cuerpo para generar el ECG.
RELACIÓN DE LA CONTRACCIÓN AURICULAR Y VENTRICULAR CON LAS ONDAS DEL ELECTROCARDIOGRAMA.
Antes de que se pueda producir la contracción del músculo, la despolarización se debe propagar por todo el músculo para iniciar los procesos químicos de la contracción. La onda P se produce al comienzo de la contracción de las aurículas y el complejo QRS de ondas se produce al comienzo de la contracción de los ventrículos. Los ventrículos siguen contraídos hasta después de que se haya producido la repolarización, es decir, hasta después del final de la onda T. Las aurículas se repolarizan aproximadamente 0,15 a 0,2 s después de la finalización de la onda P, lo que coincide aproximadamente con el momento en el que se registra el complejo QRS en el ECG. Por tanto, la onda de repolarización auricular, conocida como onda T auricular, habitualmente está oscurecida por el complejo QRS, que es mucho mayor. Por este motivo raras veces se observa la onda T auricular en el ECG. La onda de repolarización ventricular es la onda T del ECG normal. Habitualmente el músculo ventricular comienza a repolarizarse en algunas fibras aproximadamente 0,2 s después del comienzo de la onda de despolarización (el complejo QRS), pero en muchas otras fibras tarda hasta 0,35 s. Así, el proceso de repolarización ventricular se extiende a lo largo de un período prolongado, de aproximadamente 0,15 s. Por este motivo la onda T del ECG normal es una onda prolongada, aunque el voltaje de la onda T es mucho menor que el voltaje del complejo QRS, en parte debido a esta duración prolongada.
Calibración del voltaje y el tiempo del electrocardiograma.
Todos los registros de los ECG se hacen con líneas de calibración adecuadas sobre el papel de registro. Estas líneas de calibración pueden estar ya señaladas en el papel, como ocurre cuando se utiliza un registrador de pluma, o se registran en el papel al mismo tiempo que se registra el ECG, como en los tipos fotográficos de electrocardiógrafos. Las líneas de calibración horizontal están dispuestas de modo que 10 de las divisiones de las líneas pequeñas hacia arriba o hacia abajo en el ECG estándar representan 1 mV, con la positividad hacia arriba y la negatividad hacia abajo.
Las líneas verticales del ECG son las líneas de calibración del tiempo. Un ECG típico se realiza a una velocidad de papel de 25 mm/s, aunque en ocasiones se emplean velocidades más rápidas. Por tanto, cada 25 mm en dirección horizontal corresponden a 1 s y cada segmento de 5 mm, indicado por las líneas verticales oscuras, representa 0,2 s. Después los intervalos de 0,2 s están divididos en cinco intervalos más pequeños por líneas finas, cada una de las cuales representa 0,04 s.
Voltajes normales en el electrocardiograma.
Los voltajes de las ondas que se registran en el ECG normal dependen de la manera en la que se aplican los electrodos a la superficie del cuerpo y de la proximidad de los electrodos al corazón. Cuando un electrodo está colocado directamente sobre los ventrículos y un segundo electrodo está localizado en otra localización del cuerpo alejada del corazón, el voltaje del complejo QRS puede ser de hasta 3 a 4 mV. Incluso este voltaje es pequeño en comparación con el potencial de acción monofásico de 110 mV que se registra directamente en la membrana del músculo cardíaco. Cuando los ECG se registran con electrodos en los dos brazos o en un brazo y una pierna, el voltaje en el complejo QRS habitualmente es de 1 a 1,5 mV desde el punto más elevado de la onda R hasta el punto más profundo de la onda S; el voltaje de la onda P está entre 0,1 y 0,3 mV, y el de la onda T está entre 0,2 y 0,3 mV.
INTERVALO P-Q P-R
El tiempo que transcurre entre el comienzo de la onda P y el comienzo del complejo QRS es el intervalo que hay entre el inicio de la excitación eléctrica de las aurículas y el inicio de la excitación de los ventrículos. Este período se denomina intervalo P-Q. El intervalo P-Q normal es de aproximadamente 0,16 s. (Con frecuencia este intervalo se denomina intervalo P-R porque es probable que no haya onda Q.)
Intervalo Q-T
La contracción del ventrículo dura casi desde el comienzo de la onda Q (onda R si no hay onda Q) hasta el final de la onda T. Este intervalo se denomina intervalo Q-T y habitualmente es de aproximadamente 0,35 s.
Determinación de la frecuencia del latido cardíaco a partir del electrocardiograma
La frecuencia del latido cardíaco se puede determinar fácilmente a partir del ECG porque la frecuencia cardíaca es el recíproco del intervalo de tiempo entre dos latidos cardíacos sucesivos. Si el intervalo entre dos latidos, que se determina a partir de las líneas de calibración del tiempo, es de 1 s, la frecuencia cardíaca es de 60 latidos/min. El intervalo normal entre dos complejos QRS sucesivos en una persona adulta es de aproximadamente 0,83 s, lo que corresponde a una frecuencia cardíaca de 60/0,83 veces por minuto, o 72 latidos/min.
FLUJO DE CORRIENTE ALREDEDOR DEL CORAZÓN DURANTE EL CICLO CARDÍACO
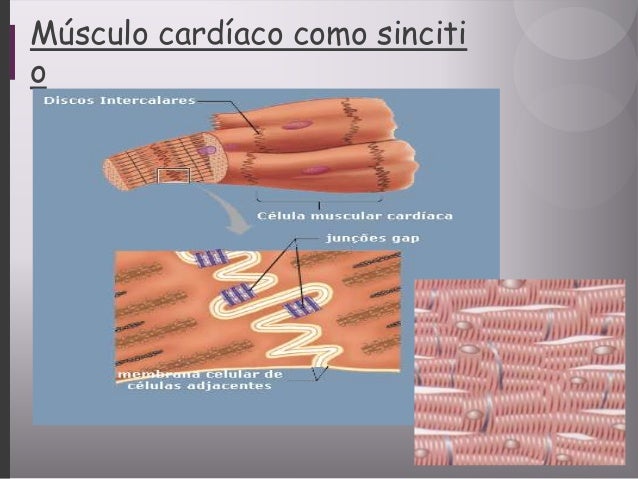
REGISTRO DE POTENCIALES ELÉCTRICOS A PARTIR DE UNA MASA PARCIALMENTE DESPOLARIZADA DE MÚSCULO CARDÍACO SINCITIAL.
Una masa sincitial de músculo cardíaco que ha sido estimulada en su punto más central. Antes de la estimulación, el exterior de todas las células musculares era positivo y el interior negativo. Por los motivos en el análisis de los potenciales de membrana, tan pronto como se despolariza una zona del sincitio cardíaco se produce la salida de cargas negativas hacia el exterior de las fibras musculares despolarizadas, haciendo que esta parte de la superficie sea electronegativa. El resto de la superficie del corazón, que sigue polarizada, está representado por los signos positivos. Por tanto, un medidor conectado con el terminal negativo en la zona de despolarización y el terminal positivo en una de las zonas que todavía están polarizadas, como se muestra a la derecha de la figura, registra un valor positivo.
También se presentan otras dos disposiciones de los electrodos y lecturas de los medidores. Estas disposiciones y lecturas se deben estudiar cuidadosamente, y el lector debe ser capaz de explicar las causas de las respectivas lecturas de los medidores. Como la despolarización se propaga por el corazón en todas las direcciones, las diferencias de potencial persisten solo durante algunas milésimas de segundo, y las mediciones del voltaje real solo se pueden realizar con un aparato de registro de alta velocidad.
FLUJO DE CORRIENTES ELÉCTRICAS EN EL TÓRAX ALREDEDOR DEL CORAZÓN
El músculo ventricular situado en el interior del tórax. Incluso los pulmones, aunque están llenos de aire en su mayor parte, conducen la electricidad en una magnitud sorprendente, y los líquidos de los demás tejidos que rodean el corazón conducen la electricidad incluso con más facilidad. Por tanto, el corazón realmente está suspendido en un medio conductor. Cuando una porción de los ventrículos se despolariza y, por tanto, se hace electronegativa en relación con el resto, la corriente eléctrica fluye desde la zona despolarizada hacia la zona polarizada en rutas sinuosas largas.
Del análisis del sistema de Purkinje se debe recordar que la primera zona de los ventrículos a la que llega el impulso cardíaco es el tabique, y poco después se propaga a la superficie interna del resto de la masa de los ventrículos, por las zonas rojas y los signos negativo. Este proceso hace que las zonas internas de los ventrículos sean electronegativas y que las paredes externas de los ventrículos sean electropositivas, de modo que la corriente eléctrica fluye a través de los líquidos que rodean los ventrículos en trayectos elípticos. Si se realiza el promedio algebraico de todas las líneas de flujo de corriente (las líneas elípticas) se encuentra que el flujo medio de corriente tiene negatividad hacia la base del corazón y positividad hacia la punta. Durante la mayor parte del resto del proceso de despolarización la corriente también sigue fluyendo en esta misma dirección, mientras que la despolarización se propaga desde la superficie endocárdica hacia el exterior a través de la masa del músculo ventricular. Después, inmediatamente antes de que la despolarización haya completado su trayecto a través de los ventrículos, la dirección media del flujo de corriente se invierte durante aproximadamente 0,01 s, fluyendo desde la punta ventricular hacia la base, porque la última parte del corazón que se despolariza son las paredes externas de los ventrículos cerca de la base del corazón.
Así, en los ventrículos del corazón normal la corriente fluye desde las zonas negativas a las positivas principalmente en una dirección que va desde la base del corazón hacia la punta durante casi todo el ciclo de despolarización, excepto al final. Si se conecta un medidor a los electrodos de la superficie del cuerpo, el electrodo más próximo a la base será negativo, mientras que el electrodo más próximo a la punta será positivo, y el medidor de registro mostrará un registro positivo en el ECG.
DERIVACIONES ELECTROCARDIOGRÁFICAS
Las conexiones eléctricas entre las extremidades del paciente y el electrocardiógrafo para registrar ECG de las denominadas derivaciones bipolares estándar de las extremidades. El término «bipolar» significa que el electrocardiograma se registra a partir de dos electrodos que están localizados en lados diferentes del corazón, en este caso en las extremidades. Así, una «derivación» no es un único cable que procede del cuerpo, sino una combinación de dos cables y sus electrodos para formar un circuito completo entre el cuerpo y el electrocardiógrafo. En cada uno de los casos el electrocardiógrafo se representa en el diagrama mediante un medidor eléctrico, aunque el electrocardiógrafo real es un sistema informático de alta velocidad con una pantalla electrónica.
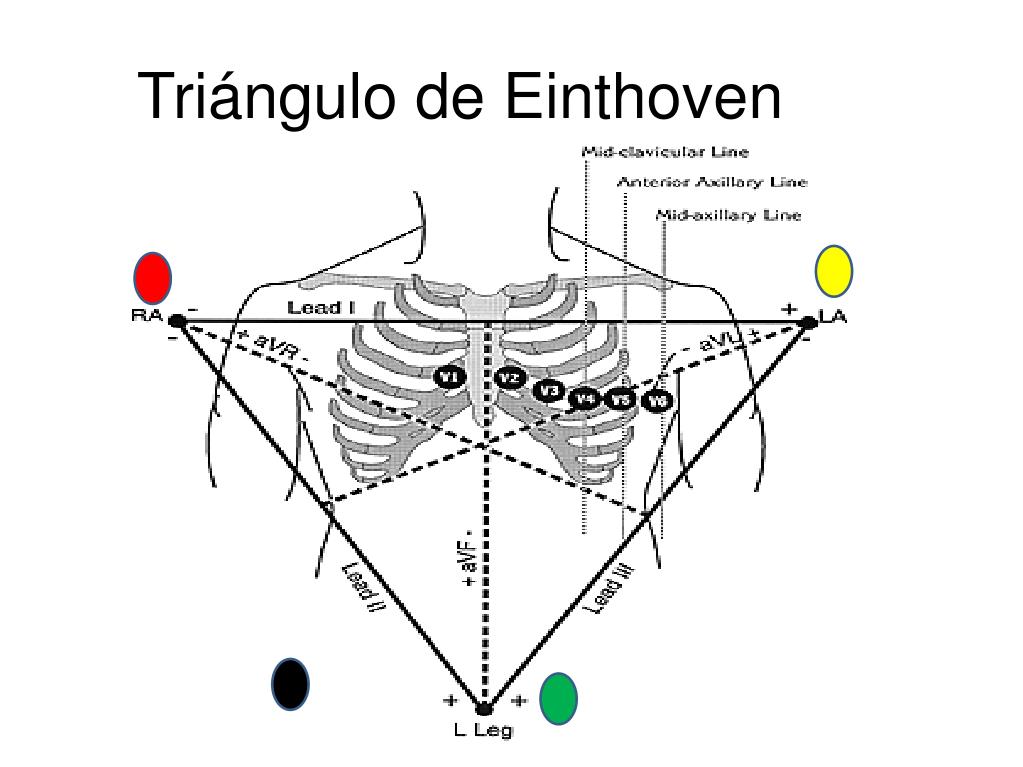
Derivación I
Cuando se registra la derivación I, el terminal negativo del electrocardiógrafo está conectado al brazo derecho y el terminal positivo al brazo izquierdo. Por tanto, cuando el punto en el que el brazo derecho se conecta con el tórax es electronegativo respecto al punto en el que se conecta el brazo izquierdo el electrocardiógrafo registra una señal positiva, es decir, por encima de la línea de voltaje cero del ECG. Cuando ocurre lo contrario el electrocardiógrafo registra una señal por debajo de la línea.
Derivación II
Para registrar la derivación II de las extremidades, el terminal negativo del electrocardiógrafo se conecta al brazo derecho y el terminal positivo a la pierna izquierda. Por tanto, cuando el brazo derecho es negativo respecto a la pierna izquierda, el electrocardiógrafo registra una señal positiva.
Derivación III
Para registrar la derivación III de las extremidades, el terminal negativo del electrocardiógrafo se conecta al brazo izquierdo y el terminal positivo a la pierna izquierda. Esta configuración significa que el electrocardiógrafo registra una señal positiva cuando el brazo izquierdo es negativo respecto a la pierna izquierda.
Triángulo de Einthoven
En el cuerpo se dibuja un triángulo, denominado triángulo de Einthoven, alrededor de la zona del corazón. Este diagrama ilustra que los dos brazos y la pierna izquierda forman vértices de un triángulo que rodea el corazón. Los dos vértices de la parte superior del triángulo representan los puntos en los que los dos brazos se conectan eléctricamente a los líquidos que rodean el corazón y el vértice izquierdo es el punto en el que la pierna izquierda se conecta a los líquidos.
Ley de Einthoven
La ley de Einthoven afirma que, si los ECG se registran simultáneamente en las tres derivaciones de las extremidades, la suma de los potenciales registrados en las derivaciones I y III debe ser igual al potencial en la derivación II.
En otras palabras, si en cualquier momento dado se conocen los potenciales eléctricos de dos cualesquiera de las tres derivaciones electrocardiográficas bipolares de las extremidades, se puede determinar la tercera simplemente sumando las dos primeras. Ha de tenerse en cuenta, sin embargo, que se deben observar los signos positivos y negativos de las diferentes derivaciones cuando se haga esta suma.
DERIVACIONES DEL TÓRAX (DERIVACIONES PRECORDIALES)
Con frecuencia se registran ECG con un electrodo situado en la superficie anterior del tórax directamente sobre el corazón en uno de los puntos. Este electrodo se conecta al terminal positivo del electrocardiógrafo, y el electrodo negativo, denominado electrodo indiferente, se conecta a través de resistencias eléctricas iguales al brazo derecho, al brazo izquierdo y a la pierna izquierda al mismo tiempo. Habitualmente se registran seis derivaciones estándar del tórax, una cada vez, desde la pared torácica anterior, de modo que el electrodo del tórax se coloca secuencialmente en los seis puntos que se muestran en el diagrama. Los diferentes registros se conocen como derivaciones V1, V2, V3, V4, V5 y V6.
En las derivaciones V1 y V2 los registros QRS del corazón normal son principalmente negativos porque, como se muestra en la figura 11-8, el electrodo del tórax de estas derivaciones está más cerca de la base del corazón que de la punta, y la base del corazón está en la dirección de la electronegatividad durante la mayor parte del proceso de despolarización ventricular. Por el contrario, los complejos QRS de las derivaciones V4, V5 y V6 son principalmente positivos porque el electrodo del tórax de estas derivaciones está más cerca de la punta cardíaca, que está en la dirección de la electropositividad durante la mayor parte de la despolarización.
Otro sistema de derivaciones que se utiliza mucho es la derivación unipolar ampliada de las extremidades. En este tipo de registro, dos de las extremidades se conectan mediante resistencias eléctricas al terminal negativo del electrocardiógrafo, y la tercera extremidad se conecta al terminal positivo. Cuando el terminal positivo está en el brazo derecho la derivación se conoce como derivación aVR, cuando está en el brazo izquierdo es la derivación aVL y cuando está en la pierna izquierda es la derivación aVF.
El corazón humano tiene un sistema especial para la autoexcitación rítmica y la contracción repetitiva aproximadamente 100.000 veces al día, o 3.000 millones de veces en una vida humana de duración media. Este impresionante logro es realizado por un sistema que:
Genera impulsos eléctricos rítmicos para iniciar la contracción rítmica del músculo cardíaco.
Conduce estos estímulos rápidamente por todo el corazón.
Cuando este sistema funciona normalmente, las aurículas se contraen aproximadamente 1/6 de segundo antes de la contracción ventricular, lo que permite el llenado de los ventrículos antes de que bombeen la sangre a través de los pulmones y de la circulación periférica. Este sistema también es importante porque permite que todas las porciones de los ventrículos se contraigan casi simultáneamente, lo que es esencial para una generación de presión más eficaz en las cavidades ventriculares.
Este sistema rítmico y de conducción del corazón se puede lesionar en las cardiopatías, especialmente en la isquemia de los tejidos cardíacos que se debe a una disminución del flujo sanguíneo coronario. La consecuencia es con frecuencia una alteración del ritmo cardíaco o una secuencia anormal de contracción de las cavidades cardíacas, con una posible alteración grave de la eficacia de la función de bomba del corazón, incluso hasta el grado de producir la muerte.
SISTEMA DE EXCITACIÓN ESPECIALIZADO Y DE CONDUCCIÓN DEL CORAZÓN
El sistema especializado de excitación y conducción del corazón que controla las contracciones cardíacas. El nódulo sinusal (también denominado nódulo sinoauricular o SA), en el que se genera el impulso rítmico normal; las vías internodulares que conducen impulsos desde el nódulo sinusal hasta el nódulo auriculoventricular (AV); el nódulo AV, en el cual los impulsos originados en las aurículas se retrasan antes de penetrar en los ventrículos; el haz AV, que conduce impulsos desde las aurículas hacia los ventrículos, y las ramas izquierda y derecha del haz de fibras de Purkinje, que conducen los impulsos cardíacos por todo el tejido de los ventrículos.
NÓDULO SINUSAL (SINOAURICULAR)
El nódulo sinusal (también denominado nódulo sinoauricular) es una banda elipsoide, aplanada y pequeña de músculo cardíaco especializado de aproximadamente 3 mm de anchura, 15 mm de longitud y 1 mm de grosor. Está localizado en la pared posterolateral superior de la aurícula derecha, inmediatamente inferior y ligeramente lateral a la desembocadura de la vena cava superior. Las fibras de este nódulo casi no tienen filamentos musculares contráctiles y cada una de ellas tiene solo de 3 a 5 μm de diámetro, en contraste con un diámetro de 10 a 15 μm para las fibras musculares auriculares circundantes. Sin embargo, las fibras del nódulo sinusal se conectan directamente con las fibras musculares auriculares, de modo que todos los potenciales de acción que comienzan en el nódulo sinusal se propagan inmediatamente hacia la pared del músculo auricular.
RITMICIDAD ELÉCTRICA AUTOMÁTICA DE LAS FIBRAS SINUSALES
Algunas fibras cardíacas tienen la capacidad de autoexcitación, que es un proceso que puede producir descargas y contracciones rítmicas automáticas. Esta capacidad es especialmente cierta en el caso de las fibras del sistema especializado de conducción del corazón, entre ellas las fibras del nódulo sinusal. Por este motivo el nódulo sinusal habitualmente controla la frecuencia del latido de todo el corazón, como se analiza en detalle más adelante en este mismo capítulo. En primer lugar, se va a describir esta ritmicidad automática.
Mecanismo de la ritmicidad del nódulo sinusal. Los potenciales de acción registrados desde el interior de una fibra del nódulo sinusal durante tres latidos cardíacos y, a modo de comparación, un único potencial de acción de una fibra muscular ventricular. Entre descargas el «potencial de membrana en reposo» de la fibra del nódulo sinusal tiene una negatividad de aproximadamente –55 a –60 mV, en comparación con –85 a – 90 mV para la fibra muscular ventricular. La causa de esta menor negatividad es que las membranas celulares de las fibras sinusales son permeables naturalmente a los iones sodio y calcio, y las cargas positivas de los iones sodio y calcio que entran neutralizan parte de la negatividad intracelular.
La ritmicidad de las fibras del nódulo sinusal muestran de que el músculo cardíaco tiene tres tipos de canales iónicos de membrana que tienen funciones importantes en la generación de los cambios de voltaje en el potencial de acción. Los tipos son:
Los canales rápidos de sodio.
Los canales de calcio de tipo L (canales lentos de sodio-calcio)
Los canales de potasio.
La apertura de los canales rápidos de sodio durante algunas diezmilésimas de segundo es responsable de la rápida espiga ascendente del potencial de acción que se observa en el músculo ventricular, debido a la entrada rápida de iones sodio positivos hacia el interior de la fibra. Después, la «meseta» del potencial de acción ventricular está producida principalmente por la apertura más lenta de los canales lentos de sodio-calcio, que dura aproximadamente 0,3 s. Finalmente, la apertura de los canales de potasio permite la difusión de grandes cantidades de iones potasio positivos hacia el exterior a través de la membrana de la fibra y devuelve el potencial de membrana a su nivel de reposo.
Hay una diferencia en la función de estos canales en la fibra del nódulo sinusal porque el potencial «de reposo» es mucho menos negativo (de solo –55 mV en la fibra nodular, en lugar de los –90 mV de la fibra muscular ventricular). A este nivel de –55 mV, los canales rápidos de sodio principalmente ya se han «inactivado», lo que significa que han sido bloqueados. La causa de esto es que siempre que el potencial de membrana es menos negativo de aproximadamente –55 mV durante más de algunos milisegundos, las compuertas de inactivación del interior de la membrana celular que cierran los canales rápidos de sodio se cierran y permanecen de esta manera. Por tanto, solo se pueden abrir los canales lentos de sodio-calcio (es decir, se pueden «activar») y, por tanto, pueden producir el potencial de acción. En consecuencia, el potencial de acción del nódulo auricular se produce más lentamente que el potencial de acción del músculo ventricular. Además, después de la producción del potencial de acción, el regreso del potencial a su estado negativo también se produce lentamente, en lugar del regreso súbito que se produce en la fibra ventricular.
Autoexcitación de las fibras del nódulo sinusal
Debido a la elevada concentración de iones sodio en el líquido extracelular en el exterior de la fibra nodular, así como al número moderado de canales de sodio abiertos previamente, los iones sodio positivos del exterior de las fibras normalmente tienden a desplazarse hacia el interior. Por tanto, entre los latidos cardíacos, la entrada de iones sodio de carga positiva produce una elevación lenta del potencial de membrana en reposo en dirección positiva. Así, el potencial «en reposo» aumenta gradualmente y se hace menos negativo entre cada dos latidos sucesivos. Cuando el potencial alcanza un voltaje umbral de aproximadamente –40 mV, los canales de calcio de tipo L se «activan», produciendo de esta manera el potencial de acción. Por tanto, básicamente, la permeabilidad inherente de las fibras del nódulo sinusal a los iones sodio y calcio produce su autoexcitación.
Durante el transcurso del potencial de acción se producen dos fenómenos que impiden dicho estado de despolarización constante. Primero, los canales de calcio de tipo L se inactivan (es decir, se cierran) en un plazo de aproximadamente 100 a 150 ms después de su apertura, y segundo, aproximadamente al mismo tiempo se abren números muy elevados de canales de potasio. Por tanto, se interrumpe el flujo de entrada de iones positivos calcio y sodio a través de los canales de calcio de tipo L, mientras que al mismo tiempo grandes cantidades de iones positivos de potasio difunden hacia el exterior de la fibra. Estos dos efectos reducen el potencial intracelular hasta devolverlo a su nivel de reposo negativo y, por tanto, ponen fin al potencial de acción. Además, los canales de potasio permanecen abiertos durante algunas décimas de segundo más, manteniendo transitoriamente el movimiento de cargas positivas hacia el exterior de la célula, con el consiguiente exceso de negatividad en el interior de la fibra; este proceso se denomina hiperpolarización. El estado de hiperpolarización inicialmente desplaza el potencial de membrana «en reposo» hacia abajo hasta aproximadamente –55 o –60 mV al final del potencial de acción. Pero, ¿por qué este nuevo estado de hiperpolarización no se mantiene indefinidamente? El motivo es que en las décimas de segundo siguientes al final del potencial de acción se cierran cada vez más canales de potasio. Los iones sodio y calcio que fluyen hacia el interior una vez más compensan el flujo de salida de iones potasio, lo que lleva a que el potencial «de reposo» se desplace hacia arriba una vez más, alcanzando finalmente el nivel liminal de aproximadamente –40 mV. Después comienza de nuevo todo el proceso: autoexcitación para generar el potencial de acción, recuperación del potencial de acción, hiperpolarización después de que haya finalizado el potencial de acción, desplazamiento del potencial «de reposo» hasta el umbral y, finalmente, reexcitación para generar un nuevo ciclo. Este proceso continúa indefinidamente durante toda la vida de una persona.
LAS VÍAS INTERNODULARES E INTERAURICULARES TRANSMITEN IMPULSOS CARDÍACOS A TRAVÉS DE LAS AURÍCULAS
Los extremos de las fibras del nódulo sinusal se conectan directamente con las fibras musculares auriculares circundantes. Por tanto, los potenciales de acción que se originan en el nódulo sinusal viajan hacia estas fibras musculares auriculares. De esta manera, el potencial de acción se propaga por toda la, masa muscular auricular y, finalmente, llega hasta el nódulo AV. La velocidad de conducción en la mayor, parte del músculo auricular es de aproximadamente 0,3 m/s, pero la conducción es más rápida, de aproximadamente 1 m/s, en varias pequeñas bandas de fibras auriculares. Una de estas bandas, denominada banda interauricular anterior, atraviesa las paredes anteriores de las aurículas para dirigirse hacia la aurícula izquierda. Además, otras tres bandas pequeñas se incurvan a través de las paredes auriculares anterior, lateral y posterior, y terminan en el nódulo AV; y se denominan, respectivamente, vías internodulares anterior, media y posterior.
La causa de la velocidad de conducción más rápida de estas bandas es la presencia de fibras de conducción especializadas. Estas fibras son similares a las «fibras de Purkinje» de los ventrículos, que conducen incluso más rápidamente.
EL NÓDULO AURICULOVENTRICULAR RETRASA LA CONDUCCIÓN DEL IMPULSO DESDE LAS AURÍCULAS A LOS VENTRÍCULOS.
El sistema de conducción auricular está organizado de modo que el impulso cardíaco no viaja desde las aurículas hacia los ventrículos demasiado rápidamente; este retraso da tiempo para que las aurículas vacíen su sangre hacia los ventrículos antes de que comience la contracción ventricular. El retraso de la transmisión hacia los ventrículos se produce principalmente en el nódulo AV y en sus fibras de conducción adyacentes.
El nódulo AV está localizado en la pared posterolateral de la aurícula derecha, inmediatamente detrás de la válvula tricúspide, muestra en forma de diagrama las diferentes partes de este nódulo, más sus conexiones con las fibras de las vías internodulares auriculares entrantes y el haz AV de salida. Esta figura también muestra los intervalos temporales aproximados en fracciones de segundo entre el comienzo inicial del impulso cardíaco en el nódulo sinusal y su posterior aparición en el sistema del nódulo AV. Obsérvese que el impulso, después de viajar por las vías internodulares, llega al nódulo AV aproximadamente 0,03 s después de su origen en el nódulo sinusal. Después hay un retraso de otros 0,09 s en el propio nódulo AV antes de que el impulso entre en la porción penetrante del haz AV, a través del cual pasa hacia los ventrículos. Se produce un retraso final de otros 0,04 s principalmente en este haz AV penetrante, que está formado por múltiples fascículos pequeños que atraviesan el tejido fibroso que separa las aurículas de los ventrículos.
Así, el retraso total en el nódulo AV y en el sistema de AV es de aproximadamente 0,13 s. Este retraso, añadido al retraso inicial de la conducción de 0,03 s desde el nódulo sinusal hasta el nódulo AV, hace que haya un retraso total de 0,16 s antes de que la señal excitadora llegue finalmente al músculo ventricular que se está contrayendo.
Causa de la conducción lenta
La conducción lenta en las fibras transicionales, nodulares y penetrantes del haz AV está producida principalmente por la disminución del número de uniones en hendidura entre células sucesivas de las vías de conducción, de modo que hay una gran resistencia a la conducción de los iones excitadores desde una fibra de conducción hasta la siguiente. Por tanto, es fácil ver por qué se tarda en excitar células sucesivas.
TRANSMISIÓN RÁPIDA EN EL SISTEMA DE PURKINJE VENTRICULAR
Las fibras de Purkinje especiales se dirigen desde el nódulo AV a través del haz AV hacia los ventrículos. Excepto en la porción inicial de estas fibras, donde penetran en la barrera fibrosa AV, tienen características funcionales bastante distintas a las de las fibras del nódulo AV. Son fibras muy grandes, incluso mayores que las fibras musculares ventriculares normales, y transmiten potenciales de acción a una velocidad de 1,5 a 4 m/s, una velocidad aproximadamente seis veces mayor que la del músculo ventricular normal y 150 veces mayor que la de algunas de las fibras del nódulo AV. Esta velocidad permite una transmisión casi instantánea del impulso cardíaco por todo el resto del músculo ventricular.
Se piensa que la rápida transmisión de los potenciales de acción por las fibras de Purkinje está producida por un gran aumento del nivel de permeabilidad de las uniones en hendidura de los discos intercalados entre las células sucesivas que componen las fibras de Purkinje. Por tanto, los iones pasan fácilmente de una célula a la siguiente, aumentando de esta manera la velocidad de la transmisión. Las fibras de Purkinje también tienen muy pocas miofibrillas, lo que significa que se contraen poco o nada durante la transmisión de los impulsos.
Conducción unidireccional a través del haz AV
Una característica especial del haz AV es la imposibilidad, excepto en estados anormales, de que los potenciales de acción viajen retrógradamente desde los ventrículos hacia las aurículas. Esta característica impide la reentrada de los impulsos cardíacos por esta ruta desde los ventrículos hacia las aurículas, permitiendo solo la contracción anterógrada desde las aurículas hacia los ventrículos.
Además, se debe recordar que en todas las localizaciones excepto en el haz AV el músculo auricular está separado del músculo ventricular por una barrera fibrosa continua. Esta barrera normalmente actúa como aislante para impedir el paso de los impulsos cardíacos entre el músculo auricular y ventricular a través de cualquier ruta distinta a la conducción anterógrada a través del propio haz AV. (En casos infrecuentes un puente muscular anormal penetra en la barrera fibrosa en otra localización distinta al haz AV. En estas condiciones el impulso cardíaco puede entrar en las aurículas desde los ventrículos y producir arritmias cardíacas graves.)
Distribución de las fibras de Purkinje en los ventrículos: las ramas izquierda y derecha del haz
Después de penetrar en el tejido fibroso que está entre el músculo auricular y ventricular, la porción distal del haz AV se dirigía hacia abajo en el interior del tabique interventricular a lo largo de 5 a 15 mm hacia la punta del corazón. Después el haz se divide en las ramas izquierda y derecha del haz, que están debajo del endocardio en los dos lados respectivos del tabique interventricular. Cada una de las ramas se dirige hacia abajo, hacia la punta del ventrículo, dividiéndose progresivamente en ramas más pequeñas. Estas ramas, a su vez, siguen un trayecto en dirección lateral alrededor de cada una de las cavidades ventriculares y hacia atrás, hacia la base del corazón. Los extremos de las fibras de Purkinje penetran en aproximadamente un tercio del grosor de la masa muscular y finalmente se continúan con las fibras musculares cardíacas.
Desde el momento en el que el impulso cardíaco entre las ramas del haz en el tabique interventricular hasta que sale de las terminaciones de las fibras de Purkinje el tiempo total transcurrido es en promedio de solo 0,03 s. Por tanto, una vez que el impulso cardíaco ha entrado en el sistema de conducción ventricular de Purkinje, se propaga casi inmediatamente a toda la masa del músculo ventricular.
TRANSMISIÓN DEL IMPULSO CARDÍACO EN EL MÚSCULO VENTRICULAR
Una vez que el impulso llega a los extremos de las fibras de Purkinje se transmite a través de la masa del músculo ventricular por las propias fibras musculares ventriculares. La velocidad de transmisión es ahora solo de 0,3 a 0,5 m/s, una sexta parte de la velocidad de las fibras de Purkinje. El músculo cardíaco envuelve el corazón en una doble espiral, con tabiques fibrosos entre las capas en espiral; por tanto, el impulso cardíaco no viaja necesariamente directamente hacia el exterior, hacia la superficie del corazón, sino que se angula hacia la superficie a lo largo de las direcciones de las espirales. Debido a esta inclinación, la transmisión desde la superficie endocárdica a la superficie, epicárdica del ventrículo precisa hasta otros 0,03 s, aproximadamente igual al tiempo necesario para la, transmisión por toda la porción ventricular del sistema de Purkinje. Así, el tiempo total para la transmisión del impulso cardíaco desde las ramas iniciales del haz hasta las últimas fibras del músculo ventricular en el corazón normal es de aproximadamente 0,06 s
CONTROL DE LA EXCITACIÓN Y LA CONDUCCIÓN EN EL CORAZÓN
EL NÓDULO SINUSAL ES EL MARCAPASOS NORMAL DEL CORAZÓN
En el análisis que se ha realizado hasta ahora sobre la génesis y transmisión del impulso cardíaco por el corazón hemos señalado que el impulso normalmente se origina en el nódulo sinusal. En algunas situaciones anormales no ocurre así. Otras partes del corazón pueden presentar también una excitación rítmica intrínseca de la misma forma que lo hacen las fibras del nódulo sinusal; esta capacidad es particularmente cierto en el caso de las fibras del nódulo AV y de las fibras de Purkinje.
Las fibras del nódulo AV, cuando no son estimuladas por alguna fuente externa, descargan a una frecuencia rítmica intrínseca de 40 a 60 veces por minuto, y las fibras de Purkinje lo hacen a una frecuencia de entre 15 y 40 veces por minuto. Estas frecuencias son distintas a la frecuencia normal del nódulo sinusal, de 70 a 80 veces por minuto.
¿Por qué entonces es el nódulo sinusal, y no el nódulo AV ni las fibras de Purkinje, el que controla la ritmicidad del corazón? La respuesta procede del hecho de que la frecuencia de descarga del nódulo sinusal es considerablemente mayor que la frecuencia de descarga autoexcitadora natural de las fibras del nódulo AV y de las fibras de Purkinje. Cada vez que se produce una descarga en el nódulo sinusal su impulso se conduce hacia el nódulo AV y hacia las fibras de Purkinje, produciendo también la descarga de sus membranas. Sin embargo, el nódulo sinusal produce una nueva descarga antes de que las fibras del nódulo AV o las fibras de Purkinje puedan alcanzar sus propios umbrales de autoexcitación. Por tanto, el nuevo impulso procedente del nódulo sinusal descarga tanto las fibras del nódulo AV como las fibras de Purkinje antes de que se pueda producir autoexcitación en cualquiera de esas estructuras.
Así, el nódulo sinusal controla el latido del corazón porque su frecuencia de descarga rítmica es más rápida que la de cualquier otra parte del corazón. Por tanto, el nódulo sinusal es casi siempre el marcapasos del corazón normal.
Marcapasos anormales: marcapasos «ectópico»
De manera ocasional alguna otra parte del corazón muestra una frecuencia de descarga rítmica que es más rápida que la del nódulo sinusal. Por ejemplo, a veces se produce este desarrollo en el nódulo AV o en las fibras de Purkinje cuando una de estas estructuras se altera. En ambos casos el marcapasos del corazón se desplaza desde el nódulo sinusal hasta el nódulo AV o las fibras de Purkinje excitadas. En casos menos frecuentes todavía, algún punto del músculo auricular o ventricular presenta una excitabilidad excesiva y se convierte en el marcapasos.
Un marcapasos que está situado en una localización distinta al nódulo sinusal se denomina marcapasos «ectópico». Un marcapasos ectópico da lugar a una secuencia anormal de contracción de las diferentes partes del corazón y puede producir una debilidad significativa del bombeo cardíaco.
Otra causa de desplazamiento del marcapasos es el bloqueo de la transmisión del impulso cardíaco desde el nódulo sinusal a las demás partes del corazón. El nuevo marcapasos se produce en este caso con más frecuencia en el nódulo AV o en la porción penetrante del haz AV en su trayecto hacia los ventrículos.
Cuando se produce un bloqueo AV, es decir, cuando el impulso cardíaco no puede pasar desde las aurículas hacia los ventrículos a través del sistema del nódulo AV y del haz, las aurículas siguen latiendo a la frecuencia normal del ritmo del nódulo sinusal, mientras que habitualmente aparece un nuevo marcapasos en el sistema de Purkinje de los ventrículos que activa el músculo ventricular a una frecuencia de entre 15 y 40 latidos/min. Después de un bloqueo súbito del haz AV el sistema de Purkinje no comienza a emitir sus impulsos rítmicos intrínsecos hasta 5 a 20 s después porque, antes del bloqueo, las fibras de Purkinje habían estado «sobreexcitadas» por los rápidos impulsos sinusales y, en consecuencia, están en un estado suprimido. Durante estos 5 a 20 s los ventrículos dejan de bombear sangre y la persona se desvanece después de los primeros 4 a 5 s debido a la ausencia de flujo sanguíneo cerebral. Este retraso de la recuperación del corazón se denomina síndrome de Stokes-Adams. Si el período de retraso es demasiado largo, se puede producir la muerte.
LOS NERVIOS SIMPÁTICOS Y PARASIMPÁTICOS CONTROLAN EL RITMO CARDÍACO Y LA CONDUCCIÓN DE IMPULSOS POR LOS NERVIOS CARDÍACOS
El corazón está inervado por nervios simpáticos y parasimpáticos. Los nervios parasimpáticos (vagos) se distribuyen principalmente a los nódulos SA y AV, en mucho menor grado al músculo de las dos aurículas y apenas directamente al músculo ventricular. Por el contrario, los nervios simpáticos se distribuyen en todas las regiones del corazón, con una intensa representación en el músculo ventricular, así como en todas las demás zonas.
La estimulación parasimpática (vagal) ralentiza el ritmo y la conducción cardíacos
La estimulación de los nervios parasimpáticos que llegan al corazón (los vagos) hace que se libere la hormona acetilcolina en las terminaciones nerviosas. Esta hormona tiene dos efectos principales sobre el corazón. Primero, reduce la frecuencia del ritmo del nódulo sinusal, y segundo, reduce la excitabilidad de las fibras de la unión AV entre la musculatura auricular y el nódulo AV, retrasando de esta manera la transmisión del impulso cardíaco hacia los ventrículos.
Una estimulación vagal débil a moderada reduce la frecuencia del bombeo del corazón, con frecuencia hasta un valor tan bajo como la mitad de lo normal. Además, la estimulación intensa de los nervios vagos puede interrumpir completamente la excitación rítmica del nódulo sinusal o puede bloquear completamente la transmisión del impulso cardíaco desde las aurículas hacia los ventrículos a través del nódulo AV. En cualquiera de los casos, las señales excitadoras rítmicas ya no se transmiten hacia los ventrículos. Los ventrículos pueden dejar de latir durante 5 a 20 s, pero después alguna área pequeña de las fibras de Purkinje, habitualmente en la porción del tabique interventricular del haz AV, presenta un ritmo propio y genera la contracción ventricular a una frecuencia de 15 a 40 latidos/min. Este fenómeno se denomina escape ventricular.
Mecanismo de los efectos vagales
La acetilcolina que se libera en las terminaciones nerviosas vagales aumenta mucho la permeabilidad de las membranas de las fibras a los iones potasio, lo que permite la salida rápida de potasio desde las fibras del sistema de conducción. Este proceso da lugar a un aumento de la negatividad en el interior de las fibras, un efecto que se denomina hiperpolarización, que hace que este tejido excitable sea mucho menos excitable.
En el nódulo sinusal, el estado de hiperpolarización hace el potencial de membrana «en reposo» de las fibras del nódulo sinusal mucho más negativo de lo habitual, es decir, de –65 a –75 mV en lugar del nivel normal de –55 a –60 mV. Por tanto, el aumento inicial del potencial de membrana del nódulo sinusal que produce la corriente de entrada de sodio y de calcio tarda mucho más en alcanzar el potencial liminal para la excitación. Este requisito retrasa mucho la frecuencia de ritmicidad de estas fibras nodulares. Si la estimulación vagal es lo suficientemente intensa es posible detener totalmente la autoexcitación rítmica de este nódulo.
En el nódulo AV, el estado de hiperpolarización producido por la estimulación vagal hace que sea difícil que las pequeñas fibras auriculares que entran en el nódulo generen una corriente de una intensidad suficiente como para excitar las fibras nodulares. Por tanto, el factor de seguridad para la transmisión del impulso cardíaco a través de las fibras de transición hacia las fibras del nódulo AV disminuye. Una reducción moderada simplemente retrasa la conducción del impulso, aunque una disminución grande bloquea totalmente la conducción.
La estimulación simpática aumenta el ritmo y la conducción del corazón
La estimulación simpática produce esencialmente los efectos contrarios sobre el corazón a los que produce la estimulación vagal. Primero, aumenta la frecuencia de descarga del nódulo sinusal. Segundo, aumenta la velocidad de conducción, así como el nivel de excitabilidad de todas las porciones del corazón. Tercero, aumenta mucho la fuerza de contracción de toda la musculatura cardíaca, tanto auricular como ventricular.
En breve, la estimulación simpática aumenta la actividad global del corazón. La estimulación máxima casi puede triplicar la frecuencia del latido cardíaco y puede aumentar la fuerza de la contracción del corazón hasta dos veces.
Mecanismo del efecto simpático
La estimulación de los nervios simpáticos libera la hormona noradrenalina en las terminaciones nerviosas simpáticas. La noradrenalina estimula, a su vez, los receptores β1-adrenérgicos, que median en los efectos sobre la frecuencia cardíaca. No está del todo claro el mecanismo preciso mediante el que la estimulación β1-adrenérgica actúa sobre las fibras del músculo cardíaco, aunque se piensa que aumenta la permeabilidad de la membrana de las fibras a los iones sodio y calcio. En el nódulo sinusal, un aumento de la permeabilidad a sodio-calcio genera un potencial en reposo más positivo y también produce un aumento de la velocidad del ascenso del potencial de membrana diastólico hacia el nivel liminal para la autoexcitación, acelerando de esta forma la autoexcitación y, por tanto, aumentando la frecuencia cardíaca.
En el nódulo AV y en los haces AV, el aumento de la permeabilidad a sodio-calcio hace que sea más fácil que el potencial de acción excite todas las porciones sucesivas de los haces de las fibras de conducción, disminuyendo de esta manera el tiempo de conducción desde las aurículas hasta los ventrículos. El aumento de la permeabilidad a los iones calcio es responsable al menos en parte del aumento de la fuerza contráctil del músculo cardíaco bajo la influencia de la estimulación simpática, porque los iones calcio tienen una función importante en la excitación del proceso contráctil de las miofibrillas.
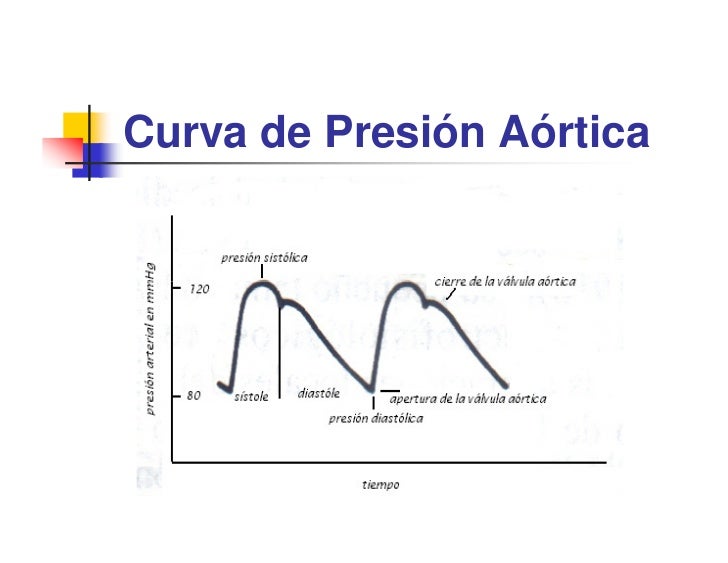
Cuando el ventrículo izquierdo se contrae, la presión ventricular aumenta rápidamente hasta que se abre la válvula aórtica. Posteriormente, después de que se haya abierto la válvula, la presión del ventrículo aumenta mucho menos rápidamente, porque la sangre sale inmediatamente del ventrículo hacia la aorta y después hacia las aterías de distribución sistémica. La entrada de sangre en las arterias hace que las paredes de las mismas se distiendan y que la presión aumente hasta aproximadamente 120 mmHg. Al final de la sístole, después de que el ventrículo izquierdo haya dejado de impulsar sangre y se haya cerrado la válvula aórtica, las paredes elásticas de las arterias mantienen una presión elevada en las arterias, incluso durante la diástole.
Se produce –> incisura en la curva de presión aórtica cuando se cierra la válvula aórtica; está producida por un corto período de flujo retrógrado de sangre inmediatamente antes del cierre de la válvula, seguido por la interrupción súbita del flujo retrógrado. Después de que se haya cerrado la válvula aórtica, la presión en el interior de la aorta disminuye lentamente durante toda la sístole porque la sangre que está almacenada en las arterias elásticas distendidas fluye contínuamente a través de los vasos periféricos de nuevo hacia las venas. Antes de que se contraiga de nuevo el ventrículo, la presión aórtica habitualmente ha disminuido hasta 80 mmHg (presión diastólica), que es dos tercios de la presión máxima de 120 mmHg (presión sistólica) que se produce durante la contracción ventricular.Las curvas de presión de ventrículo derecho y arteria pulmonar son similares, excepto que las presiones tienen un magnitud aprox. de 1/6.
RELACIÓN DE LOS TONOS CARDÍACOSCON EL BOMBEO CARDÍACO.
Cuando se contraen los ventrículos primero se oye un ruido que está producido por el cierre de las válvulas auriculoventriculares. La vibración tiene un tono bajo y es prolongada –> primer tono cardíaco. Cuando se cierran las válvulas aórtica y pulmonar al final de la sístole se oye un golpe seco y rápido porque estas válvulas se cierran rápidamente y los líquidos circundantes vibran durante un período corto –> segundo tono cardíaco.
GENERACIÓN DE TRABAJO DEL CORAZÓN.
El trabajo sistólico del corazón –> cantidad de energía que el corazón convierte en trabajo durante cada latido cardíaco mientras bombea sangre hacia las arterias. El trabajo minuto –> cantidad total de energía que se convierte en trabajo en un minuto (= trabajo sistólico x frecuencia cardíaca por minuto).El trabajo del corazón se utiliza de dos maneras:
1) La mayor proporción se utiliza para mover la sangre desde las venas de baja presión hacia las arterias de alta presión (trabajo volumen- presión o trabajo externo).
2) Una pequeña proporción de la energía se utiliza para acelerar la sangre hasta su velocidad de eyección a través de las válvulas aórtica y pulmonar. Este es el componente de energía cinética del flujo sanguíneo del trabajo cardíaco.
El trabajo externo del ventrículo derecho es la 1/6 del izquierdo, debido la diferencia de 6 veces de las presiones sistólicas que bombean los dos ventrículos. El trabajo adicional para generar energía cinética del flujo sanguíneo es proporcional a la masa de sangre que se expulsa, por la velocidad de eyección al cuadrado –> = m. v
Análisis gráfico del bombeo ventricular.
La curva de presión diastólica se determina llenando el corazón con volúmenes de sangre progresivamente mayores y midiendo la presión diastólica inmediatamente antes de que se produzca la contracción ventricular, que es la presión tele diastólica del ventrículo.La curva de presión sistólica se determina registrando la presión sistólica que se alcanza durante la contracción ventricular a cada volumen llenado.Hasta que el volumen del ventrículo que no se está contrayendo no aumenta por encima de aproximadamente 150 ml, la presión diastólica no aumenta mucho. Por tanto, hasta este volumen la sangre puede fluir con facilidad hacia el ventrículo desde la aurícula. Por encima de 150 ml la presión diastólica ventricular aumenta rápidamente, en parte porque el tejido fibroso del corazón ya no se puede distender más y en parte porque el pericardio que rodea el corazón se ha llenado casi hasta su límite.Durante la contracción ventricular, la presión sistólica aumenta a volúmenes ventriculares bajos y alcanza un máximo a un volumen ventricular de 150 a 170ml. Después a medida que sigue aumentando el volumen, la presión sistólica llega a disminuir en algunas situaciones porque a estos volúmenes elevados los filamentos de actina y de miosina de las fibras musculares cardíacas están tan separados que la fuerza de la contracción de cada una de las fibras cardíacas se hace menos óptima.
Presión sistólica normal ventrículo izquierdo –> 250- 300 mmHg.
Presión sistólica normal ventrículo derecho –> 60-80 mmHg
Diagrama volumen presión durante el ciclo cardíaco. Trabajo cardíaco.
Fase 1: período de llenado (línea I) : comienza a un volumen ventricular de aproximadamente 45 ml y una presión diastólica próxima a 0 mmHg. 45 ml es la can+dad de sangre que queda en el ventrículo después del la+do previo(volumen tele sistólico – precarga). A medida que la sangre venosa fluye hacia el ventrículo desde la aurícula izquierda, el volumen ventricular aumenta hasta 115 ml (volumen tele diastólico). La presión diastólica aumenta hasta 5 mmHg.
Fase 2: período de contracción isovolúmica (línea II) : el volumen del ventrículo no se modifica porque todas las válvulas están cerradas. La presión en el interior del ventrículo aumenta hasta igualarse a la presión que hay en la aorta, hasta aproximadamente 80 mmHg.
Fase 3: período de eyección (línea III) : la presión sistólica aumenta más debido a la contracción aún más intensa del ventrículo. El volumen del ventrículo disminuye porque la válvula aórtica ya se ha abierto y la sangre sale del ventrículo hacia la aorta.
Fase 4: período de relajación isovolúmica (línea IV) : al final del período de eyección se cierra la válvula aórtica, y la presión ventricular disminuye de nuevo hasta el nivel de la presión diastólica. Así el ventrículo recupera su valor inicial, en el que quedan aproximadamente 45 ml de sangre en el ventrículo y la presión auricular es de 0 mmHg.
Concepto de precarga y poscarga
Precarga: grado de tensión del músculo cuando comienza a contraerse.
Poscarga: carga contra la que el músculo ejerce su fuerza contráción.
Para la contracción cardíaca se considera que la precarga es la presión tele diastólica cuando el ventrículo ya se ha llenado. La poscarga del ventrículo es la presión de la arteria que sale del ventrículo, la presión que se necesita para que la sangre sea expulsada de ventrículo a arteria (presión sistólica).
ENERGÍA QUÍMICA NECESARIA PARA LA CONTRACCIÓN CARDÍACA: LA UTILIZACIÓN DE OXÍGENO POR EL CORAZÓN.
Músculo cardíaco: utliza energía química para realizar el trabajo de la contracción. La velocidad del consumo de O2 por el miocardio es una medida excelente de la energía química que se libera mientras el corazón realiza su trabajo.Durante la contracción la mayor parte de la energía química que se gasta se convierte en calor y una porción mucho menor en trabajo. El cociente del trabajo respecto al gasto de energía total se denomina eficiencia de la contracción cardíaco o eficiencia del corazón. La eficiencia máxima normal está entre el 20-25%.
REGULACIÓN DEL BOMBEO CARDÍACO
Cuando una persona está en reposo el corazón solo bombea de 4 a6 litros de sangre cada minuto. Durante el ejercicio intenso puede ser necesario que el corazón bombee de 4 a 7 veces esta cantidad. Los mecanismos básicos mediante los que se regula el volumen que bombea el corazón son:
Regulación cardíaca intrínseca del bombeo en respuesta a los cambios del volumen de la sangre que fluye hacia el corazón.
Control de la frecuencia cardíaca y del bombeo cardíaco por el SNA.
Regulación intrínseca del bombeo cardíaco: el mecanismo de Frank- Starling.
La cantidad de sangre que bombea el corazón cada minuto está determinada casi totalmente por la velocidad el flujo sanguíneo hacia el corazón desde las venas (retorno venoso). Todos los tejidos periféricos del cuerpo controlan su propio Bujo sanguíneo local, y todos los flujos tisulares locales se combinan y regresan a través de las venas hacia la aurícula derecha. El corazón a su vez bombea automáticamente hacia las arterias esta sangre que le llega, de modo que pueda fluir de nuevo por el circuito.Esta capacidad intrínseca del corazón de adaptarse a volúmenes crecientes de flujo sanguíneo de entrada –> mecanismo de Frank Starling del corazón–> cuanto más se distiende el músculo cardíaco durante el llenado, mayor es la fuerza de contracción y mayor es la cantidad de sangre que bombea hacia la aorta (el corazón bombea toda la sangre que le llega procedente de las venas).
Efecto de los iones potasio y calcio sobre la función cardíaca.
Exceso de K: hace que el corazón este dilatado y Bácido, y también reduce la frecuencia cardíaca. Grandes cantidades también pueden bloquear la conducción del impulso desde las aurículas hacia los ventrículos a través del haz auriculoventricular. La elevación de la concentración a 8 a 12 mEq/L puede provocar la muerte. (valor normal 3-4mEq/L)
Exceso de Ca: efectos casi contrarios a los de K, haciendo que el corazón progrese hacia una contracción espástica por el efecto directo de los Ca en el inicio de la contracción (déficit de Ca: Bacidez cardíaca)
EXCITACIÓN RÍTMICA DEL CORAZÓN
El corazón está dotado de un sistema especial para:
Generar impulsos eléctricos rítmicos para producir la contracción rítmica del músculo cardíaco.
Conducir estos estímulos rápidamente por todo el corazón.
Cuando este sistema funciona normalmente las aurículas se contraen aproximadamente 1/6 de segundo, antes de la contracción ventricular, lo que permite el llenado de los ventrículos antes de que bombeen la sangre a través de los pulmones y de la circulación periférica. Este sistema también es importante porque permite que todas las porciones de los ventrículos se contraigan casi simultáneamente lo que es esencial para una generación de presión más eficaz en las cavidades ventriculares.
SISTEMA DE EXCITACIÓN ESPECIALIZADO Y DE CONDUCCIÓN DEL CORAZÓN
Nódulo sinusal: donde se genera el impulso cardiaco normal
Vías internodulares: conducen el impulso desde nódulo sinusal hasta nódulo auriculoventricular.
Nódulo auriculoventricular (AV): impulso se retrasa antes de penetrar en los ventrículos.
Haz AV: conduce el impulso desde las aurículas hasta los ventrículos
Ramas izquierdas y derechas de bras de Purkinje: conducen el impulso por todo el tejido de los ventrículos.
1) Nódulo sinusal: banda elipsoide, aplanada y pequeña de músculos cardíaco especializado. Localizado en la pared posterolateral superior de la aurícula derecha, inmediatamente inferior y lateral a la desembocadura de la VCS. Las fibras de este, casi no tienen filamentos contráctiles. Sin embargo, estas se conectan directamente con las fibras musculares auriculares de modo que todos los potenciales de acción que comienzan en él se propagan hacia la pared del músculo auricular.
2) Vías internodulares y transmisión del impulso cardíaco a través de las aurículas: potenciales de acción viajan de nódulo sinusal hacia estas fibras musculares auriculares circundantes. El potencial de acción se propaga por toda la masas auricular y finalmente llega al nódulo auriculoventricular. La velocidad de conducción en la mayor parte del músculo auricular es de aproximadamente 0,3 m/s, pero la conducción es más rápida, de aproximadamente 1 m/s en varias bandas de fibras auriculares. Una de ellas –> banda interauricular anterior: atraviesa las paredes anteriores de las aurículas para dirigirse hacia la aurícula izquierda. Otras 3 bandas pequeñas se incurvan a través de las paredes auriculares anterior, lateral y posterior y terminan en el nódulo auriculoventricularvías internodulares anterior, media y posterior. La causa de la velocidad de conducción más rápida de estas bandas es la presencia de fibras de conducción especializada, similar a las fibras de Purkinje de los ventrículos.
3) Nódulo auriculoventricular: el retraso de la transmisión hacia los ventrículos (para que aurículas se vacíen) se produce principalmente aquí y en las fibras de conducción adyacentes. El nódulo está localizado en la pared posterolateral de la aurícula derecha, detrás de la válvula tricúspide. El impulso después de viajar por las vías internodulares, llega al nódulo auriculoventricular aproximadamente 0,03 segundos después de su origen en el nódulo sinusal. Después hay un retraso de 0,09 seg. en el propio nódulo AV antes de que el impulso entre en la porción penetrante del haz auriculoventricular, a través del cual pasa hacia los ventrículos. Se produce un retraso 2nal de 0,04 segundos formado por múltiples fascículos pequeños que atraviesan el tejido fibroso que separa las aurículas de los ventrículos. El retraso total es de 0,16 segundos antes de que la señal excitadora llegue finalmente al músculo ventricular que se está contrayendo. El retraso del nódulo y del sistema AV es de 0,13 seg y se le añade 0,03seg desde el nódulo sinusal hasta el AV. Causa de la conducción lenta disminución del número de uniones de hendidura entre células sucesivas de las vías de conducción. Hay una gran resistencia a la conducción de los iones excitadores desde una fibra de conducción a la siguiente.
4) Sistema de Purkinje: las fibras de Purkinje se dirigen desde el nódulo AV a través del haz AV hacia los ventrículos. Son fibras muy grandes y transmiten potenciales de acción a una velocidad de 1,5 a 4 m/s, una velocidad aproximadamente 6 veces mayor que la del músculo ventricular normal y 150 veces más que las fibras nodulares AV. Esto permite una transmisión casi instantánea del impulso por todo el resto del músculo ventricular. La rápida transmisión está producida por un gran aumento del nivel de permeabilidad de las uniones en hendidura de los discos intercalados entre las células sucesivas que componen las fibras de Purkinje –> los iones pasan fácilmente de una célula a la siguiente, aumentando la velocidad de la transmisión. Las fibras de Purkinje también tienen muy pocas miofibrillas, lo que significa que se contraen poco o nada durante la transmisión de impulsos.Una característica del haz de His, es la imposibilidad de que los potenciales de acción viajen retrógradamente desde los ventrículos hacia las aurículas. Además, se debe recordar que en todas las localizaciones, excepto en el haz, el musculo auricular está separado del ventricular por una barrera fibrosa continua que actúa como aislante para impedir el paso de los impulsos entre estos músculos a través de cualquier ruta distinta a la conducción anterógrada a través del haz de His.
5) Transmisión del impulso cardíaco en el músculo ventricular: la velocidad de transmisión ahora es de 0,3 a 0,5 m/s. El músculo cardíaco envuelve el corazón en una doble espiral, con tabiques fibrosos entre las capas en espiral; el impulso cardíaco no viaja directamente hacia el exterior (hacia la superficie del corazón) sino que se angula a la superficie a lo largo de las direcciones de las espirales. El tiempo total para la transmisión del impulso desde las ramas iniciales del haz hasta las últimas fibras del músculo ventricular es de aprox. 0,06 seg.
CONTROL DE LA EXCITACIÓN Y LA CONDUCCIÓN EN EL CORAZÓN
El nódulo sinusal como marcapasos del corazón. La frecuencia de descarga del nódulo sinusal es mayor que la frecuencia de descarga auto excitadora natural de las fibras del nódulo auriculoventricular y de las fibras de Purkinje. Cada vez que se produce una descarga en el nódulo sinusal su impulso se conduce hacia el nódulo auriculoventricular y hacia las fibras de Purkinje, produciendo también la descarga de sus membranas. Pero el nódulo sinusal produce una nueva descarga antes de que las fibras del nódulo auriculoventricular o las fibras de Purkinje puedan alcanzar sus propios umbrales de autoexcitación. Por tanto, el nuevo impulso procedente del nódulo sinusal descarga tanto las fibras del nódulo auriculoventricular como las fibras de Purkinje antes de que se pueda producir autoexcitación en cualquiera de sus estructuras.Así el nódulo sinusal controla el la+do del corazón porque su frecuencia de descarga rítmica es más rápida que la de cualquier otra parte del corazón el nódulo sinusal es prácticamente siempre el marcapasos del corazón normal.
Importancia del sistema de Purkinje en la generación de una contracción sincrónica del músculo ventricular
Normalmente el impulso cardíaco llega a casi todas las porciones de los ventrículos en un breve intervalo de tiempo, excitando la primera fibra muscular ventricular solo 0,03 a 0,06 segundos antes de la excitación de la última. Esto hace que todas las porciones del músculo de los dos ventrículos comiencen a contraerse casi al mismo tiempo y que después sigan contrayéndose durante aproximadamente otros 0,3 segundos. La función de bomba eficaz de las dos cavidades ventriculares precisa este tipo sincrónico de contracción. Si el impulso cardíaco viajara lentamente a través de los ventrículos, buena parte de la masa ventricular se contraería antes de la contracción del resto, en cuyo caso se produciría una gran disminución de la función global de bomba.
Control del ritmo cardíaco y la conducción de impulsos por los nervios cardíacos
Nervios parasimpáticos se distribuyen principalmente:
Nódulo SA.
Nódulo AV.
↓músculo auricular ↓↓ músculo ventricular.
Nervios simpáticos se distribuyen principalmente:
Todas las regiones del corazón.
Representación intensa en músculo ventricular.
La estimulación parasimpática (vagal) y escape ventricular.
La estimulación de los nervios parasimpáticos hace que se libere acetilcolina en las terminaciones nerviosas. Esta hormona tiene dos efectos principales sobre el corazón:
Reduce la frecuencia del ritmo del nódulo sinusal.
Reduce la excitabilidad de las fibras de la unión auriculoventricular entre la musculatura auricular y el nódulo auriculoventricular, retrasando de esta manera la transmisión del impulso cardíaco hacia los ventrículos.
Una estimulación vagal débil a moderada reduce la frecuencia del bombeo del corazón, con frecuencia hasta un valor tan bajo como la mitad de lo normal. Una estimulación intensa de los nervios vagos, puede interrumpir completamente la excitación rítmica del nódulo sinusal o puede bloquear completamente la transmisión del impulso cardíaco desde las aurículas hacia los ventrículos a través del nódulo auriculoventricular: señales no se transmiten a los ventrículos, los ventrículos dejan de latir durante 5 a 20 segundos pero después algún punto de las fibras de Purkinje: genera contracción ventricular a una frecuencia de 15 a 40 la+dos por minuto: escape ventricular.Mecanismo de los efectos vagales. Terminaciones nerviosas vagales –> acetulcolina aumenta permeabilidad de las membranas de K –> salida rápida de K desde las fibras al sistema de conducción–> aumento de la negatividad en interior de las fibras –> Hiperpolarización –> hace que tejido excitable sea mucho menos excitable.
Nódulo sinusal en Hiperpolarización –> reduce el potencial de membrana en reposo –> aumento inicial de potencial de membrana que produce la corriente de entrada de Na y Ca tarda mucho más en alcanzar el potencial liminal para la excitación.
Nódulo AV en Hiperpolarización –> hace que sea difícil que las pequeñas fibras auriculares que entran al nódulo generen una corriente de suficiente intensidad como para excitar las fibras nodulares.
Efecto de la estimulación simpática sobre el ritmo y la contracción del corazón
La estimulación simpática produce esencialmente los efectos contrarios sobre el corazón a los que produce la estimulación vagal:
Aumenta la frecuencia de descarga del nódulo sinusal.
Aumenta la velocidad de conducción.
Aumenta el nivel de excitabilidad de todas las porciones del corazón.
Aumenta mucho la fuerza de contracción de toda la musculatura cardíaca, tanto auricular como ventricular.
Una estimulación máxima puede triplicar la frecuencia cardíaca y doblar la contracción.
Mecanismo de efecto simpático
Nervios simpáticos –> noradrenalina –> aumenta la permeabilidad de la membrana de las fibras a Na y Ca –> genera un potencial de acción más positivo –> acelera de esta forma la autoexcitación. En el nódulo y haces AV, el aumento de la permeabilidad hace que sea más fácil que el PA excite todas las porciones sucesivas de los haces de las fibras de conducción, disminuyendo el tiempo de conducción desde aurículas a ventrículos. El aumento a la permeabilidad del Ca es responsable en parte del aumento de la fuerza contráctil porque tienen función en la excitación del proceso contráctil de las miofibrillas.
El corazón está formado por dos bombas separadas: un corazón derecha que bomba sangre hacia los pulmones y uno izquierdo, hacia los órganos periféricos. Cada uno es una bomba bicameral pulsátil formada por una aurícula y un ventrículo. Cada aurícula es una bomba débil de cebado del ventrículo que contribuye a transportar sangre al ventrículo correspondiente. Estos, aportan la principal fuerza de bombeo que impulsa la sangre:
Hacia la circulación pulmonar por el ventrículo derecho
Hacia la circulación periférica por el ventrículo izquierdo.
Mecanismos especiales producen una sucesión continuada de contracciones cardíaca denominada, ritmicidad cardíaca; que transmite potenciales de acción por todo el musculo cardíaco y determina su latido rítmico.
Fisiología del musculo cardiaco
El corazón está formado por 3 tipos principales de musculo cardíaco:
Musculo auricular y ventricular: se contrae de manera similar al esquelético pero su duración de contracción es mucho mayor.
Fibras especializadas de contracción y de conducción: se contraen solo débilmente ya que tienen pocas fibras contráctiles. Presentan descargas eléctricas rítmicas automáticas, en forma de potenciales de acción o conducción de los potenciales de acción por todo el corazón.
Anatomía Fisiológica del musculo cardiaco.
Las fibras musculares cardíacas están dispuestas en un retículo de modo que se dividen, se vuelven a combinar y se separan de nuevo. Es estriado. Posee miofibrillas típicas que contienen filamentos de actina y miosina. Se encuentran uno al lado del otro y se deslizan entre sí, durante la contracción, de la misma manera que el musculo esquelético.
Musculo esquelético como sincitio. Las fibras musculares cardíacas están formadas por muchas células individuales conectadas entre sí, en serie y en paralelo. Los discos intercalares son membranas celulares que separan estas células musculares cardíacas individuales entre sí. Las membranas se fusionan de tal manera que forman uniones comunicantes en hendidura, permeables, que permiten una difusión de iones casi totalmente libre. Por tanto, se mueven con facilidad en el líquido intracelular, a lo largo del eje longitudinal de las fibras; así los potenciales de acción viajan fácilmente desde una célula muscular cardíaca a la siguiente y el musculo cardíaco funciona como un sincitio. El corazón está formado por dos sincitios: el auricular y el ventricular. Las aurículas, están separadas de los ventrículos por tejido fibroso que rodea las aberturas de las válvulas auriculoventriculares. Normalmente los potenciales no se conducen desde el sincitio auricular hacia el ventricular directo, a causa del tejido fibroso. Sino que son conducidos por medio de un sistema de conducción especializado denominado haz auriculoventricular; que es un fascículo de fibras de conducción de varios mm de diámetro. La división en dos sincitios permite que las aurículas se contraigan un pequeño intervalo antes que la contracción ventricular lo que es importante para la eficacia del bombeo del corazón.
Potenciales de acción (PA) en el musculo cardiaco. El PA que se registra en una fibra muscular ventricular es en promedio de aprox 105mV, lo que significa que el potencial intracelular aumenta desde un valor muy negativo de -85mV entre los la+dos, hasta un valor ligeramente positivo de +20mV. Después de la espiga inicial la membrana permanece despolarizada durante aproximadamente 0,2 segundos mostrando una meseta, seguida al final de esta de una repolarización súbita. La presenta de esta meseta hace que la contracción ventricular dura hasta 15 veces más en el musculo cardíaco que en el musculo esquelético.
¿Qué produce el PA prolongado y la meseta? Son responsables de esto, dos diferencias importantes entre las propiedades de la membrana del musculo cardíaco y esquelético.
1) El PQ del musculo esquelético, esta producido casi por completo por la apertura súbita de grandes números de los canales de sodio que permiten que grandes cantidades de sodio entren en la fibra. A estos se los denomina canales rápidos, porque permanecen abiertos solo algunas milésimas de segundos y luego se cierran súbitamente. Al final de este cierre, se produce la repolarización y el PA ha terminado en otra milésima de seg aprox. En el musculo cardíaco, el PA está producido por la apertura de dos +pos de canales: 1. Los mismos canales rápidos de sodio q en el musculo esquelético y 2. Otros canales totalmente distintos: canales lentos de calcio, también llamados canales de calcio-sodio; estos difieren de los canales rápidos de sodio ya que se abren con mayor lentitud y permanecen abiertos durante varias décimas de segundos. Durante este tiempo, fluyen gran cantidad de iones calcio y sodio hacia el interior manteniendo un periodo prolongado de despolarización, dando lugar a la meseta del PA.
2) Inmediatamente después del inicio del PA, la permeabilidad de la membrana del musculo cardíaco a los iones potasio, disminuye aproximadamente 5 veces. Efecto que no aparece en el musculo esquelético. Esta disminución de la permeabilidad, se puede deber al exceso de flujo de entrada de calcio a través de los canales de calcio. Independientemente de la causa, la disminución de la permeabilidad reduce mucho el flujo de salida de iones potasio de carga positiva durante la meseta del potencial de acción, impidiendo el regreso rápido del voltaje de potencial de membrana a su nivel de reposo. Cuando los canales lentos de calcio-sodio se cierran después de 0,2 a 0,3 segundos y se interrumpe el flujo de entrada de estos, también aumenta rápidamente la permeabilidad a los iones potasio, esta rápida perdida de potasio devuelve el potencial de membrana a su nivel de reposo.
Velocidad de la conducción de la señales en el musculo cardíaco: es de aproximadamente 0,3 a 0,5 m/s en las fibras auriculares y ventriculares. La velocidad de conducción en el sistema especializado de conducción del corazón, en las fibras de Purkinje, es de hasta 4 m/s; lo que permite una conducción rápida de las señales excitadores hacia las diferentes partes del corazón.
Periodo refractario. El musculo cardíaco es refractario a la reestilmulacion durante el PA. El período refractario del corazón, es el periodo de tiempo durante el cual un impulso cardíaco normal, no puede reexitar una zona ya excitada del musculo. La duración normal en el ventrículo es de 0,25 a 0,3 segundos. Hay un periodo refractario relativo adicional de aprox 0,05 segundos, durante el cual es más diFcil de lo normal excitar el musculo. Sin embargo, se puede excitar con una señal excitadora muy intensa causando una extrasístole temprana.
Duración de la contracción: el musculo comienza a contraerse pocos milisegundos después de la llegada del PA, y sigue contrayéndose algunos segundos después de que finalice. Por lo tanto, depende de la duración del PA: 0,2 seg en el musculo auricular y 0,3 en el ventricular.
Ciclo cardiaco.
Fenómenos cardíacos que se producen desde el comienzo de un la+do hasta el comienzo del siguiente. Cada ciclo es iniciado por la generación espontanea de un PA en el nódulo sinusal, localizado en la pared supero lateral de la aurícula derecha, cerca del orificio de la vena cava superior. El PA viaja por ambas aurículas y después a través de las AV hacia los ventrículos. Debido a esta disposición, hay un retraso de más de 0,1 seg durante el paso del impulso cardíaco desde las aurículas a los ventrículos, permitiendo bombear sangre hacia los ventrículos antes que comience su contracción.
Relación del electrocardiograma con el ciclo cardíaco. El electrocardiograma muestras las ondas P, Q, R, S y T. Son, los voltajes eléctricos que genera el corazón. La onda P, está producida por la propagación de la despolarización en las aurículas yes seguida por la contracción auricular que produce una ligera elevación de la curva de presión auricular, inmediatamente después de la onda P. Aproximadamente 0,16 segundos del inicio de la onda P, la ondas Q, R y S aparecen como consecuencia de la despolarización de los ventrículos que inician su contracción y hace que comience a elevarse la presión ventricular. Así, el complejo QRS comienza un poco antes del inicio de la sístole ventricular. Finalmente, se observa la onda T, ventricular, que representa la fase de repolarización de los ventrículos cuando las fibras del musculo ventricular comienza a relajarse. Por lo tanto, se produce un poco antes que del final de la contracción ventricular.
Funcion de los ventrículos como bombas.
Llenado de ventrículos: durante la sístole ventricular se acumulan grandes cantidades de sangre en las aurículas, porque las válvulas AV están cerradas. Cuando finaliza la sístole, y las presiones ventriculares disminuyen a los valores diastólicos, el aumento de presión que se ha generado en las aurículas abren las válvulas AV y permiten que la sangre fluya hacia los ventrículos. Esto se denomina periodo de llenado rápido: dura aprox el primer tercio de la diástole. Durante el tercio medio, solo fluye una pequeña can+dad de sangre hacia los ventrículos; esta es la sangre que continua drenando hacia las aurículas desde las venas y que pasa directamente hacia los ventrículos. Durante el último tercio, las aurículas se contraen y aportan un impulso adicional al flujo de entrada de sangre hacia los ventrículos.
Vaciado de los ventrículos durante la sístole.
Periodo de contracción isométrica. Después del comienzo de la contracción ventricular, se produce un aumento súbito de presión ventricular que hace que las válvulas AV se cierren. Después, son necesarios otros 0,2 a 0,3 seg para que el ventrículo acumule una presión suficiente para abrir las válvulas AV semilunares contra las presiones de la aorta y de la arteria pulmonar. Por lo tanto, en este periodo se produce la contracción pero no el vaciado de los ventrículos.
Periodo de eyección. Cuando la presión ventricular izquierda aumenta ligeramente por encima de 80mm Hg y la presión derecha por encima de los 8mm Hg, las presiones ventriculares abren las válvulas semilunares y comienza a salir sangre de modo que el 70% del vaciado se produce durante el primer tercio del periodo de eyección (periodo de eyección rápida) y el 30% durante los dos tercios siguientes (periodo de eyección lenta).
Periodo de relajación isométrica. Al final de la sístole, comienza súbitamente la relajación ventricular, permitiendo que las presiones disminuyan rápidamente. Las presiones elevadas de las grandes arterias que se acaban de llenar desangre, empujan la sangre inmediatamente de nuevo hacia los ventrículos lo que cierran las válvulas aorticas y pulmonares. Durante otros 0,3 a 0,6 el musculo continúa relajándose aun cuando no se modifica el volumen ventricular. Durante este periodo las presiones disminuyen rápidamente y regresan a sus valores diastólicos. Después se abren las válvulas AV para comenzar un nuevo ciclo.
Volumen tele diastólico, volumen tele sistólico y volumen sistólico.
Volumen tele diastólico: durante la diástole, el llenado de los ventrículos aumenta su volumen aprox 110 a 120 ml.
Volumen sistólico: a medida que se vacían durante la sístole el volumen disminuye a 70ml. Volumen tele sistólico: volumen restante, aprox 40 a 50ml. La fracción del volumen tele diastólico que es propulsada, se denomina fracción de eyección (aprox el 60%). Cuando el corazón se contrae con fuerza, el volumen tele sistólico puede disminuir hasta 10 o 20 ml, por el contrario cuando Buyen grandes cantidades hacia los ventrículos durante la diástole, los volúmenes tele diastólicos pueden llegar a 150 o 180ml. El aumento del volumen tele diastólico y reducción del tele sistólico puede aumentar el volumen sistólico hasta el más del doble.
Función de las válvulas.
Válvulas auriculoventriculares (tricúspide y mitral)
Impiden el flujo retrógrado de sangre desde los ventrículos hacia las aurículas durante la sístole y las válvulas semilunares (aórtica y de arteria pulmonar) impiden el flujo retrógrado desde las arterias aorta y pulmonar hacia los ventrículos durante la diástole. Estas válvulas, se cierran y abren pasivamente. Se cierran cuando un gradiente de presión retrógrada empuje la sangre hacia atrás y se abren cuando un gradiente de presión anterógrada fuerza la sangre en dirección anterógrada.
Válvulas auriculoventriculares –> formadas por una película delgada, casi no precisan ningún flujo retrógrado para cerrarse.
Válvulas semilunares –> son mucho más fuertes, precisan un flujo retrógrado bastante rápido durante algunos milisegundos.
Función de los músculos papilares. Se unen a los velos de las válvulas AV mediante las cuerdas tendinosas. Los músculos se contraen cuando se contraen las paredes ventriculares, pero, no contribuyen al cierre de las válvulas. Por el contrario, tiran de los velos de las válvulas hacia dentro, hacia los ventrículos, para impedir que protruyan demasiado hacia las aurículas durante la contracción ventricular.
Válvula aórtica y de la arteria pulmonar // válvulas auriculoventriculares.
Válvulas semilunares
Se cierran súbitamente por las elevadas presiones de las arterias al final de la sístole.
Debido a sus orificios más pequeños, la velocidad de la eyección de la sangre a través de ellas es mayor.
Debido al cierre rápido y a la eyección rápida, sus bordes están sometidos a una abrasión mecánica mayor.
No tienen el soporte de las cuerdas tendinosas.
Válvulas auriculoventriculares
Cierre suave.
Debido a sus orificios más grandes, la velocidad de eyección de la sangre a través de ellas es menor.
Los rayos de luz viajan a través del aire a una velocidad de unos 300.000 km/s, pero se desplazan con mucha mayor lentitud cuando recorren sólidos y líquidos transparentes.
El índice de refracción de una sustancia transparente es el cociente entre la velocidad de la luz en el aire y su velocidad en ese medio. El valor que toma en el aire es de 1.
Las lentes y la refracción
Una lente convexa concentra los rayos de luz.
Una lente cóncava dispersa los rayos de luz.
Una lente cilíndrica desvía los rayos de luz en un solo plano: comparación con las lentes esféricas.
La combinación de dos lentes cilíndricas en ángulo recto equivale a una lente esférica.
Dioptría:
Cuanto más amplia sea la desviación de los rayos luminosos por una lente, mayor es su «poder dióptrico» o poder de refracción. Este poder dióptrico se mide en dioptrías.
Óptica del ojo:
El ojo, equivale a una cámara fotográfica corriente desde el punto de vista óptico. Posee un sistema de lentes, un sistema de apertura variable (la pupila) y una retina que corresponde a la película.
El sistema ocular de lentes está compuesto por cuatro superficies de refracción: 1) la separación entre el aire y la cara anterior de la córnea 2) la separación entre la caraposterior de la córnea y el humor acuoso 3) la separación entre el humor acuoso y la cara anterior del cristalino. 4) la separación entre la cara posterior del cristalino y el humor vítreo.
El índice de refracción para el aire es 1; el de la córnea; el del humor acuoso; el del cristalino (como promedio), y el del humor vítreo.
Formación de una imagen en la retina.
De la misma manera que una lente de vidrio es capaz de enfocar una imagen sobre una hoja de papel, el sistema ocular de lentes puede enfocar una imagen sobre la retina. El resultado está dado la vuelta e invertido con respecto al objeto. Sin embargo, la mente percibe los objetos en su posición derecha a pesar de su orientación al revés en la retina debido a que el cerebro está entrenado para considerar como normal una imagen invertida.
Mecanismo de acomodación.
En los niños, el poder dióptrico del cristalino puede aumentar a voluntad desde 20 dioptrías hasta unas 34, lo que corresponde a una «acomodación» de 14 dioptrías. Para conseguirlo, su forma cambia desde una lente con una convexidad moderada hasta una lente muy convexa.
En una persona joven, el cristalino está compuesto por una potente cápsula elástica rellena de un líquido viscoso de carácter proteináceo, pero transparente. Cuando se encuentra en estado de relajación, sin ninguna tensión aplicada sobre la cápsula, adopta una forma casi esférica, debido básicamente a la retracción elástica de este elemento. Sin embargo, unos 70 ligamentos suspensorios se fijan radialmente en torno al cristalino, y tiran de sus extremos hacia el perímetro exterior del globo ocular. Estos ligamentos se encuentran constantemente tensos por sus inserciones en los bordes anteriores de la coroides y de la retina. Esta situación hace que el cristalino permanezca relativamente plano si el ojo está en condiciones normales.
La acomodación está controlada por nervios parasimpáticos.
El músculo ciliar está controlado casi en su integridad por señales nerviosas parasimpáticas transmitidas hacia el ojo desde el núcleo del tercer par en el tronco del encéfalo a través de este nervio.
La estimulación de los nervios parasimpáticos contrae los dos tipos de fibras que componen el músculo ciliar, lo que relaja los ligamentos del cristalino y propicia un aumento del grosor y del poder dióptrico de dicha estructura.
Presbicia: pérdida de acomodación en el cristalino.
A medida que una persona envejece, el cristalino crece y se engruesa perdiendo mucha elasticidad, en parte debido a la desnaturalización progresiva de sus proteínas. La capacidad que posee de modificar su forma disminuye con la edad. El poder de acomodación desciende de unas 14 dioptrías en un niño hasta menos de 2 para la época en que una persona llega a los 45 o 50 años y desciende prácticamente hasta 0 dioptrías con 70 años de edad. A partir de entonces, el cristalino queda casi totalmente desprovisto de su capacidad de acomodación, situación que se conoce como presbicia.
Errores de refracción.
Emetropía (visión normal): El ojo se considera normal, o «emétrope», si los rayos de luz paralelos procedentes de objetos alejados quedan enfocados con nitidez en la retina cuando el músculo ciliar esté relajado por completo. Esto significa que el ojo emétrope es capaz de ver todos los objetos distantes con claridad mientras el músculo ciliar se mantiene relajado.
Hipermetropía: La hipermetropía suele deberse a la presencia de un globo ocular demasiado corto o, a veces, de un sistema de lentes demasiado débil. Para vencer esta anomalía, el músculo ciliar ha de contraerse con el fin de incrementar la potencia del cristalino. Una persona hipermétrope, cuando recurre al mecanismo de la acomodación, es capaz de enfocar los objetos alejados en la retina.
Miopía: En la miopía, cuando el músculo ciliar está relajado del todo, los rayos de luz procedentes de objetos alejados quedan enfocados delante de la retina.
Astigmatismo: El astigmatismo es un error de la refracción ocular que hace que la imagen visual de un plano quede enfocada a una distancia diferente de la que corresponde a su plano perpendicular. Lo más frecuente es que el astigmatismo obedezca a una curvatura de la córnea demasiado grande en uno de los planos del ojo.
Sistema humoral del ojo: líquido intraocular.
El ojo está relleno de líquido intraocular, que mantiene una presión suficiente en el globo ocular para que siga estando dilatado. Este líquido puede dividirse en dos componentes: el humor acuoso, que se halla delante del cristalino, y el humor vítreo, que está entre la cara posterior del cristalino y la retina. El humor acuoso es un líquido que circula con libertad, mientras que el humor vítreo, a veces denominado cuerpo vítreo, es una masa gelatinosa cuya cohesión se mantiene por una fina red fibrilar compuesta básicamente por moléculas de proteoglucanos muy largas.
El «glaucoma» provoca una alta presión intraocular y es una causa fundamental de ceguera.
El glaucoma, una de las causas más frecuentes de ceguera, es una enfermedad ocular en la que asciende la presión intraocular hasta un nivel patológico, subiendo en ocasiones bruscamente hasta 60 o 70 mmHg. Las presiones por encima de 25 a 30 mmHg pueden provocar una pérdida de visión si se mantienen durante un período prolongado.
Unas presiones altísimas son capaces de ocasionar la ceguera en un plazo de días o incluso de horas. Cuando sube la presión, los axones del nervio óptico quedan comprimidos en su salida del globo ocular a través del disco óptico. Se cree que esta compresión bloquea el flujo axónico del citoplasma desde los somas neuronales situados en la retina hacia las fibras del nervio óptico que se dirigen al cerebro.
El resultado es una ausencia de la nutrición pertinente para las fibras, lo que a la larga produce la muerte de las que se vean afectadas.
Función receptora y nerviosa de la retina.
La retina es la porción del ojo sensible a la luz que contiene: 1) los conos, responsables de la visión de los colores 2) los bastones, que pueden detectar luz tenue y están encargados básicamente de la visión en blanco y negro y de la visión en la oscuridad.
Los componentes funcionales de la retina que se disponen en las siguientes capas desde el exterior hacia el interior:
1) Capa pigmentaria. 2) Capa de conos y bastones que aloja las prolongaciones de estos receptores hacia la capa anterior. 3) Capa nuclear externa que contiene los somas de los conos y los bastones. 4) Capa plexiforme externa. 5) Capa nuclear interna. 6) Capa plexiforme interna. 7) Capa ganglionar. 8) Capa de las fibras del nervio óptico. 9) Membrana limitante interna.
Conos y bastones.
Los bastones son más estrechos y largos, pero esto no siempre es así. En las porciones periféricas de la retina, los bastones alcanzan un diámetro de 2 a 5 μm, mientras que los conos miden de 5 a 8; en la parte central de la retina, la fóvea, hay bastones, y los conos son delgados y su diámetro solo es de 1,5 μm.
Capa pigmentaria de la retina.
El pigmento negro melanina de la capa pigmentaria impide la reflexión lumínica por toda la esfera del globo ocular, lo que resulta importantísimo para una visión nítida. El pigmento cumple la misma función en el ojo que el color negro en el interior de los fuelles de una cámara. Sin él, los rayos de luz se reflejarían en todas las direcciones dentro del globo y generarían una iluminación difusa de la retina en vez del contraste normal entre los puntos claros y oscuros necesario para la formación de una imagen precisa.
Desprendimiento de retina.
La porción nerviosa de la retina a veces se desprende del epitelio pigmentario. En algunos casos, la causa de este desprendimiento es una lesión del globo ocular que permite la acumulación de líquido o de sangre entre ambos elementos, la retina nerviosa y el epitelio pigmentario. En otras ocasiones, el desprendimiento está ocasionado por una contractura de las fibrillas delgadas de colágeno presentes en el humor vítreo, que tiran de zonas de la retina hacia el interior del globo.
Fotoquímica de la visión.
Tanto los conos como los bastones contienen productos químicos que se descomponen ante la exposición a la luz y, en el curso del proceso, excitan a las fibras nerviosas que salen del ojo. La sustancia sensible a la luz en los bastones se llama rodopsina; en los conos, donde se denominan pigmentos de los conos o pigmentos del color, su composición no es nada más que un poco diferente a la que presenta la rodopsina.
Ceguera nocturna.
La ceguera nocturna o hesperanopía aparece en personas con hipovitaminosis A grave ya que, sin vitamina A, la cantidad de retinal y de rodopsina que puede formarse disminuye acusadamente. Esta situación se denomina ceguera nocturna porque la magnitud de luz existente por la noche es demasiado escasa como para permitir una visión suficiente en las personas aquejadas de una deficiencia de vitamina A. Para que surja una ceguera nocturna, las personas habitualmente deben seguir una alimentación desprovista de vitamina A durante meses.
Regulación automática de la sensibilidad retiniana: adaptación a la luz y a la oscuridad.
Si una persona ha estado expuesta a una luz radiante durante horas, una gran parte de las sustancias fotosensibles en los conos y los bastones habrá quedado reducida a retinal y opsinas. Por añadidura, gran parte del retinal presente en ambos tipos de receptores se habrá convertido en vitamina A. Debido a estos dos efectos, las concentraciones de productos fotosensibles que quedan en los conos y los bastones son considerablemente bajas, y la sensibilidad del ojo a la luz está reducida de forma proporcional. Este proceso se llama adaptación a la luz.
Por el contrario, si una persona permanece en la oscuridad durante un período prolongado, el retinal y las opsinas de los conos y los bastones han vuelto a convertirse en los pigmentos sensibles a la luz. Por ende, la vitamina A se transforma de nuevo en retinal para aumentar los pigmentos fotosensibles, quedando fijado el límite final en función de la cantidad de opsinas presente en los conos y los bastones preparada para combinarse con el retinal. Este proceso se denomina adaptación a la oscuridad.
Daltonismo rojo-verde.
Cuando en el ojo solo falta un grupo de conos receptores del color, la persona es incapaz de distinguir algunos colores de otros. los colores verde, amarillo, naranja y rojo, que son los que figuran entre las longitudes de onda de 525 a 675 nm, normalmente se diferencian entre sí mediante los conos rojo y verde. Si no existe cualquiera de estos dos conos, la persona no está en condiciones de recurrir a este mecanismo para discernir entre los cuatro colores; sobre todo fallará en la distinción entre el rojo y el verde y, por tanto, se dice que tiene un daltonismo rojo-verde.
Función nerviosa de la retina.
Los diversos tipos neuronales son los siguientes:
Los fotorreceptores, los conos y los bastones, que transmiten las señales hacia la capa plexiforme externa, donde hacen sinapsis con las células bipolares y horizontales.
Las células horizontales, que transmiten las señales en sentido horizontal por la capa plexiforme externa desde los conos y los bastones hasta las células bipolares.
Las células bipolares, que transmiten las señales en sentido vertical desde los conos, los bastones y las células horizontales hacia la capa plexiforme interna, donde hacen sinapsis con las células ganglionares y amacrinas.
Las células amacrinas, que transmiten las señales en dos direcciones, directamente desde las células bipolares hasta las células ganglionares, u horizontalmente en el seno de la capa plexiforme interna desde los axones de las células bipolares hasta las dendritas de las células ganglionares o hasta otras células amacrinas.
Las células ganglionares, que transmiten las señales de salida desde la retina hacia el cerebro a través del nervio óptico.
Células amacrinas y sus funciones:
Se han identificado unas 30 clases de células amacrinas por medios morfológicos e histoquímicos. Ya se ha podido caracterizar la función más o menos de una docena de ellas y todas son diferentes.
Un tipo de célula amacrina forma parte de la vía directa para la visión de los bastones, a saber, la compuesta por el bastón, las células bipolares, las células amacrinas y las células ganglionares.
Otro tipo de célula amacrina responde potentemente cuando comienza una señal visual continua, pero su actividad se extingue con rapidez.
Otras células amacrinas presentan una respuesta enérgica al desaparecer las señales visuales, pero, una vez más, su descarga cesa a gran velocidad.
Otro tipo responde cuando se enciende o se apaga una luz, para indicar simplemente un cambio de iluminación, sin tener en cuenta su sentido.
Otro tipo más responde al movimiento de un punto a través de la retina en una dirección específica; por tanto, se dice que estas células amacrinas son sensibles a la dirección.
Así pues, y en un cierto sentido, muchas de las células amacrinas o su mayoría son interneuronas que sirven para analizar las señales visuales antes de que lleguen a abandonar la retina.
Neurofisiología central de la visión.
Vías visuales: Las principales vías visuales que van desde las dos retinas hasta la corteza visual. Las señales nerviosas de este carácter abandonan la retina a través de los nervios ópticos. En el quiasma óptico, las fibras procedentes de la mitad nasal de la retina cruzan hacia el lado opuesto, donde se unen a las fibras originadas en la retina temporal contraria para formar los tractos ópticos o cintillas ópticas. A continuación, las fibras de cada tracto óptico hacen sinapsis en el núcleo geniculado lateral dorsal del tálamo, y desde allí, las fibras geniculocalcarinas se dirigen a través de la radiación óptica (también denominada tracto geniculocalcarino) hacia la corteza visual primaria en el área correspondiente a la cisura calcarina del lóbulo occipital medial.
Las fibras visuales también se dirigen a otras regiones más antiguas del encéfalo: 1) desde los tractos ópticos llega hasta el núcleo supraquiasmático del hipotálamo, se supone que para regular los ritmos circadianos que sincronizan los diversos cambios fisiológicos del organismo según la noche y el día 2) hacia los núcleos pretectales en el mesencéfalo, para suscitar movimientos reflejos de los ojos a fin de enfocarlos sobre los objetos de importancia y activar el reflejo fotomotor pupilar 3) hacia el colículo superior, para controlar los movimientos direccionales rápidos de ambos ojos 4) hacia el núcleo geniculado lateral ventral del tálamo y las regiones basales adyacentes del cerebro, se cree que para contribuir al dominio de algunas de las funciones conductuales que lleva a cabo el organismo.
Anomalías del campo visual.
A veces, los puntos ciegos se encuentran en porciones del campo visual diferentes al área del disco óptico. Tales puntos, llamados escotomas, a menudo están causados por el daño del nervio óptico como consecuencia de un glaucoma (presión excesiva del líquido contenido en el interior del globo ocular), las reacciones alérgicas a nivel de la retina o procesos tóxicos, como el saturnismo o el consumo excesivo de tabaco.
Otro trastorno que puede diagnosticarse con la campimetría es la retinitis pigmentaria. En esta enfermedad, ciertas porciones de la retina degeneran, y en las zonas degeneradas se deposita un exceso del pigmento melanina. La retinitis pigmentaria primero suele ocasionar una ceguera en el campo visual periférico y a continuación invadir gradualmente las áreas centrales.
Campos visuales (campimetría).
El campo visual es la zona de visión observada por un ojo en un instante dado. La región percibida por el lado nasal se llama campo visual nasal y la que llega al lado lateral campo visual temporal.
Para diagnosticar una ceguera en una porción específica de la retina, se cartografía el campo visual de cada ojo mediante un procedimiento llamado campimetría. Esta cartografía se realiza haciendo que el individuo mire con un ojo hacia un punto central situado directamente delante de él; el otro ojo está cerrado.
Efecto de las lesiones de la vía óptica sobre los campos visuales.
La destrucción de todo el nervio óptico origina una ceguera en el ojo afectado. Si el trastorno afecta al quiasma óptico impide el cruce de los impulsos que proceden de la mitad nasal de cada retina y van dirigidos hacia el tracto óptico del lado opuesto. Por tanto, esta mitad queda ciega a ambos lados, lo que significa que la persona está ciega en el campo temporal de cada ojo porque la imagen del campo visual se encuentra invertida en la retina debido al sistema óptico ocular; esta situación se denomina hemianopsia bitemporal. Tales lesiones normalmente son el resultado de un tumor hipofisario que presiona sobre la parte inferior del quiasma desde la silla turca hacia arriba.
Control muscular de los movimientos oculares.
Los movimientos oculares están controlados por tres pares de músculos:
1) los rectos medial y lateral 2) los rectos superior e inferior 3) los oblicuos superior e inferior. Los rectos medial y lateral se contraen para desplazar los ojos de un lado a otro.
os rectos superior e inferior lo hacen para moverlos hacia arriba y hacia abajo. En cuanto a los músculos oblicuos, intervienen sobre todo en la rotación de los globos oculares a fin de mantener los campos visuales en posición vertical.
Control del diámetro pupilar.
La estimulación de los nervios parasimpáticos también excita el músculo esfínter de la pupila, lo que disminuye por esta vía la apertura pupilar; este proceso se denomina miosis. A la inversa, la estimulación de los nervios simpáticos excita las fibras radiales del iris y provoca la dilatación pupilar, lo que se denomina midriasis.
Reflejo pupilar fotomotor.
Cuando la luz ilumina los ojos, las pupilas se contraen, reacción llamada reflejo pupilar fotomotor. La función del reflejo fotomotor consiste en ayudar al ojo a adaptarse de forma rapidísima a unas condiciones lumínicas cambiantes. El diámetro pupilar tiene unos límites en torno a 1,5 mm por su extremo inferior y a 8 mm por el superior. Por tanto, dado que el brillo de la luz que llega a la retina crece con el cuadrado de esta variable, la amplitud de la adaptación a la luz y a la oscuridad que puede alcanzarse mediante el reflejo pupilar más o menos es de 30 a 1; es decir, la cantidad de luz que penetra en el ojo cambia hasta 30 veces.
Síndrome de Horner.
Los nervios simpáticos del ojo a veces quedan interrumpidos. Esta circunstancia suele suceder en la cadena simpática cervical, lo que provoca el cuadro clínico llamado síndrome de Horner. Este síndrome ocasiona los siguientes efectos:
Primero, debido a la interrupción de las fibras nerviosas simpáticas dirigidas al músculo dilatador de la pupila, esta permanece contraída de forma continua con un diámetro más pequeño que la pupila del lado opuesto. Segundo, el párpado superior se cae debido a que normalmente se mantiene en posición abierta durante las horas de vigilia en parte por la contracción de las fibras musculares lisas contenidas en su interior e inervadas por el sistema simpático. Por tanto, la destrucción de estos nervios imposibilita su apertura hasta una altura normal. Tercero, los vasos sanguíneos del lado correspondiente de la cara y de la cabeza quedan dilatados de un modo persistente. Cuarto, no puede producirse la sudoración (que requiere la acción de las señales nerviosas simpáticas) en el mismo lado de la cara y de la cabeza afectado por el síndrome de Horner.
Es sobre todo una función de las
yemas gustativas de la boca.
La textura y presencia de ciertas
sustancias modifican la experiencia gustativa.
La importancia del gusto es que permite
a la persona escoger la comida en función de sus deseos o necesidades
metabólicas.
SENSACIONES GUSTATIVAS PRIMARIAS
Se han identificado un mínimo de
13 receptores químicos
posibles o probables en las
células gustativas, de los siguientes tipos:
2 para el sodio
2 para el potasio
1 para el cloruro
2 para el sabor dulce
2 para el sabor amargo
1 para el glutamato
1 para el ion hidrógeno
1 para la adenosina
1 para la iosina
Se han reunido en 5 categorías
generales llamadas Sensaciones gustativas
primarias. Estas son: agrio, dulce, salado, amargo y “umami”.
Sabor AGRIO
Es causado por los ácidos y la
intensidad de esta sensación gustativa es aproximadamente proporcional al
logaritmo de esta concentración del ion hidrógeno.
Cuánto más ácido sea un alimento,
más potente se vuelve dicha sensación.
Sabor SALADO
Se despierta por las sales ionizadas,
especialmente por la concentración del ion sodio.
Los cationes de las sales son los
responsables del gusto salado, sobre todo de los cationes sodio.
Sabor DULCE
No está dado por una sola clase de
sustancia química sino en su mayoría por algunas sustancias que en su mayoría
son de origen orgánico.
Una pequeña modificación en la
estructura química como la incorporación de un radical puede cambiar el sabor
de dulce a amargo.
Sabor AMARGO
Al igual que el sabor dulce no
está dado por un determinado agente químico y en su mayoría son de origen
orgánico. Dos clases en especial pueden causar este sabor amargo:
Las
sustancias de cadena larga que contienen Nitrógeno.
Los
alcaloides: la quinina, la cafeína, la estricnina y la nicotina.
Algunas sustancias que al principio
saben saladas dejan un regusto amargo, esto sucede con la sacarina.
Sabor UMAMI
Algunos fisiólogos los consideran
el quinto sabor.
Se le denomina a una sensación
gustativa agradable, deliciosa; es sabor dominante de los alimentos que contienen
L-glutamato. Sin embargo aún no están claros los mecanismos moleculares exactos
responsables del sabor Umami.
UMBRAL GUSTATIVO
El umbral gustativo cumple una
función protectora importante contra muchas toxinas peligrosas de los
alimentos.
Ceguera gustativa
Algunas personas están ciegas
para el gusto de ciertas sustancias, sobre todo los diversos tipos de tiourea.
De todas las personas de un 15% a
30% exhiben una ceguera gustativa, el porcentaje exacto depende del método de exploración
y de la concentración de feniltiocarbamida que utilicen.
YEMAS GUSTATIVAS y su función
La yema gustativa está constituida
por 50 células epiteliales modificadas, algunas de las cuales son células de
soporte llamadas células de sostén y otras son células gustativas (sometidas a reposición continua por división mitótica).
Los extremos externos de las células
están dispuestas en torno a un Poro gustativo.
Desde este punto, sobresalen varias
microvellosidades o cilios gustativos que se dirigen hacia la cavidad oral en
el poro gustativo. Estas microvellosidades proporcionan la superficie receptora
para el gusto. Entretejida alrededor de los cuerpos de las células gustativas
hay toda una red de fibras nerviosas gustativas que reciben el estímulo de las
células receptoras del gusto. Debajo de la membrana celular se forman muchas
vesículas que se cree tienen una sustancia neurotransmisora que se libera para excitar
a las terminaciones de las fibras nerviosas como respuesta a la estimulación
gustativa
Localización de las yemas
gustativas
Se encuentran en los 3 tipos
siguientes de papilas linguales:
Una
gran cantidad está en las paredes de las depresiones que rodean a las papilas caliciformes.
Un
número moderado queda sobre las papilas fungiformes.
Una
porción también moderada se encuentran en las papilas foliáceas.
Existen otras yemas gustativas
más en el paladar y unas pocas en los pilares amigdalinos, en la epiglotis e
incluso en la parte proximal del esófago.
Adultos: de 3000 a 10000 yemas
gustativas.
Niños: tienen unas pocas más.
Pasados los 45 años muchas yemas
se degeneran.
Especificidad de las yemas gustativas
para un estímulo gustativo primario
Estudios han demostrado colocando
un microelectrodo independiente en cada yema gustativa la capacidad de responder
a cada uno de los 5 estímulos si la concentración es baja pero si la
concentración es alta puede excitarse por 2 o más estímulos que no encajan
dentro de las “categorías primarias”.
Mecanismo de estimulación de las
yemas gustativas
La membrana de la célula
gustativa tiene una carga negativa en su interior con respecto al exterior, la aplicación
de una sustancia con sabor sobre los cilios gustativos provoca una pérdida de
este potencial, es decir, se despolariza; este cambio del potencial eléctrico se
denomina potencial del receptor para el gusto. El tipo de proteína receptora en
cada vellosidad gustativa determina el tipo de gusto que vaya a percibirse.
Transmisión de las señales
gustativas en el sistema nervioso central
Los impulsos procedentes de:
LOS 2/3 ANTERIORES DE LA LENGUA
Se dirigen primero al N. Lingual,
a continuación van por la cuerda del tímpano hacia el N. Facial y finalmente
llegan al Tracto solitario en el tronco del encéfalo.
REGIONES POSTERIORES DE LA LENGUA
Y GARGANTA
Se transmiten a través del N.
Glosofaríngeo también hacia el Tracto solitario pero a un nivel un poco más
inferior.
DESDE LA BASE DE LA LENGUA Y
REGIONES FARÍNGEAS
Por medio del N. Vago estas
señales gustativas se conducen al Tracto solitario.
Todas las fibras gustativas hacen
sinapsis en los núcleos del tracto solitario, situados en la región posterior
del tronco del encéfalo
Desde el tracto solitario muchas
señales se envían hacia los núcleos salivales inferior y superior para
controlar la secreción de saliva durante la ingestión y digestión de comida.
La adaptación del gusto se trata
de un fenómeno diferente del que se da en la mayor parte de los demás sistemas sensitivos,
cuya adaptación se da en su integridad a nivel de los receptores.
Preferencias gustativas y control
del régimen alimentario
Las preferencias gustativas no
significan nada más que un animal elegirá ciertos tipos de comida por encima de
otros, en función de las necesidades corporales de ciertas sustancias específicas.
Una razón importante para pensar que la preferencia gustativa consiste en un
fenómeno propio del sistema nervioso central radica en que las experiencias
acumuladas con sabores agradables y desagradables cumplen un cometido
importante para determinar las preferencias gustativas de cada uno.
Ejemplo: si una persona se pone
enferma por comer un determinado alimento en concreto va a desarrollar una preferencia
gustativa negativa por ese alimento en particular.
En el
oído medio, la
membrana timpánica se
fija el manubrio del
martillo y este
hueso en su
extremo se articula con el yunque
y se mantienen unidos por varios ligamentos. En el extremo medial del yunque se
articula con la cabeza del estribo y la base de este último hueso descansa sobre
el laberinto membranoso
de la cóclea en la abertura de la ventana oval. El músculo
tensor del tímpano
mantiene tensa la membrana
timpánica, el cual
permite que las vibraciones sonoras de cualquier parte de
la membrana se transmitan a los huesecillos. Los ligamentos
que suspenden a los huesecillos permiten que estos se muevan como
una sola palanca y la articulación del yunque con el estribo hace que este
empuje la ventana oval y el líquido coclear hacia dentro.
Ajuste de impedancias
Impedancia
sonora→ Es la resistencia que opone un medio a las ondas que se propagan
sobre este, es decir una forma de disipación de energía de las ondas que se
desplazan en un medio.
El aire no ejerce la fuerza
necesaria para producir vibraciones en el líquido coclear, la membrana
timpánica y el sistema de palanca de los
huesecillos reduce la distancia recorrida por el estribo, pero incrementa
la fuerza de
empuje 1,3 veces,
aportando un ajuste
de impedancias entre
las ondas sonoras y el líquido alrededor de un 50-75%. Esto permite que
se utilice la mayor parte de la energía portada por las ondas sonoras entrantes
para producir las vibraciones en el líquido. Si no hubiera este sistema, el
sonido sería casi imperceptible a un nivel de volumen medio.
Atenuación del sonido mediante la contracción de los músculos:
Estapedio y tensor del tímpano.
Reflejo de atenuación→ Cuando la cadena de huesecillos transmite
sonidos fuertes hacia el SNC, se produce la contracción de los músculos
Estapedio y tensor del tímpano, haciendo que la cadena de huesecillos
se torne rígido
y reduciendo así
enormemente la conducción
sonora de baja frecuencia. Esto cumple 3 funciones:
Proteger
la cóclea de vibraciones lascivas provocadas por sonidos excesivamente
fuertes
En
mascara sonidos de baja frecuencia en ambientes ruidosos
Disminuir
la sensibilidad auditiva frente a la propia voz
Transmisión del sonido
a través del
hueso→ Las vibraciones
del cráneo pueden
provocar vibraciones del líquido coclear, sin embrago la energía aérea
no permite oír a través del hueso,
excepto cuando se aplica directamente sobre el hueso un dispositivo
electromecánico.
Cóclea
Anatomía funcional de la cóclea→ Consta de 3 tubos enrollados; unidos por sus lados
Rampa vestibular
Rampa media o conducto coclear
Rampa timpánica
Membrana de
Reissner→ Separa la rampa
vestibular y media, es delgada y se
mueve fácilmente. No obstruye
el paso de vibraciones a
través del líquido
desde la rampa vestibular hasta
la media, ambas se consideran como una
sola cámara. Laimportancia de
esta membrana es el
mantenimiento de la endolinfa en la rampa media, necesaria
para función de células
ciliadas receptoras.
Lámina
Basilar→ Separa la
rampa timpánica y media,
sobre ella descansa
el órgano de Corti.
Es fibrosa, formada
por fibras basilares que se unen al modiolo (centro óseo de la
cóclea) y tienen un extremo libre insertado
en la membrana
basilar. A medida
que se aproximan
al helicotrema la longitud
de las fibras
va aumentado, en
cambio el diámetro
de fibras y
su rigidez disminuyen. Como
consecuencia la Resonancia
de membrana basilar
para altas frecuencias tiene
lugar cerca de la base (más rígido) y la resonancia para bajas frecuencias,
cerca del helicotrema (son menos rígidas y poseen mayor cantidad de líquido).
Transmisión de las ondas
sonoras en la cóclea (onda viajera)
Cuando el estribo se desplaza
hacia dentro contra la ventana oval, la onda de líquido viaja a través de la
lámina Basilar hacia el helicotrema
Patrón de vibración
de la lámina
basilar para las
distintas frecuencias sonoras
Las ondas
sonoras de alta
frecuencia recorren una
distancia corta por la
membrana basilar antes de llegar a su punto de resonancia y extinguirse Ondas
sonoras de frecuencia media viajan aproximadamente la mitad del recorrido y
luego desaparecen. Onda sonora de baja frecuencia recorre toda la distancia de
la membrana basilar. Onda viajera se
propaga rápidamente por
porción inicial de
membrana basilar, pero luego
progresivamente más despacio
a medida que
avanza por la cóclea.
Esto se debe
a que coeficiente
de elasticidad de
fibras basilares va disminuyendo progresivamente. Patrón de la amplitud de vibración→ Es la medida en al que
vibra la membrana basilar durante un ciclo vibratorio completo.
Función del órgano de Corti→ Órgano receptor que genera
impulsos nerviosos en respuesta a la vibración de la membrana basilar. Descansa
sobre la membrana basilar. Posee 2 tipos de Receptores especializados:
Las
células ciliadas internas (1 fila)
Las
células ciliadas externas (4 filas)
La base y lados de células ciliadas hacen sinapsis con terminaciones nerviosas cocleares (el 90-95% con las células ciliadas internas) Estas fibras llegan al ganglio espiral de Corti, el cual envía señales al nervio coclear y luego al SNC a nivel de la parte superior del bulbo.
Excitación de las células ciliadas
Los estereocilios (cilios que sobresalen desde los extremos de las células ciliadas) se proyectan desde células ciliadas y se introducen en la membrana tectoria (situada por encima del esterocilios) Inclinación de cilios en una dirección despolarizan células ciliadas y su inclinación en dirección contraria las hiperpolariza. Estos movimientos excitan a las fibras del nervio coclear. Los cilios se encuentran anclados a la lámina reticular, esta es una estructura rígida en la que están acumuladas terminaciones nerviosas de células ciliadas. Esta lámina esta sostenida por los pilares de Corti y a su vez los pilares se insertan en fibras basilares de membrana basilar. Al vibrar la membrana basilar, los pilares de Corti y láminas reticulares se desplazan también como una unidad. Este movimiento hace que los cilios reboten atrás y adelante contra la membrana.
Ajuste del
sistema receptor→ Células
ciliadas externas controlan
la sensibilidad de las
células ciliadas internas a los diferentes tonos sonoros.
Potenciales de receptor de las células ciliadas y excitación de las
fibras nerviosas auditivas
El movimiento de los cilios
provoca el desplazamiento de iones K+ desde el líquido del conducto coclear
adyacente hacia los estereocilios, y esto suscita la despolarización de la
membrana de la célula ciliada. Cuando
fibras basilares se
inclinan hacia la
rampa vestibular, células
ciliadas se despolarizan,
y cuando se mueven
en sentido contrario
se hiperpolarizan, por
lo que generan
un potencial de receptor alternante.
Potencial
endococlear→ Es el potencial eléctrico de +80 mV que existe entre la
endolinfa y la perilinfa. Es generado por transporte continuo de K+ hacia la
rampa media a través de la estría vascular.
Endolinfa→
Se encuentra dentro de la rampa media (secretada por la estría vascular)
Perilinfa→
Se encuentra dentro de la rampa vestibular y timpánica (casi idéntico al
LCR) Parte superior de células ciliadas
están bañadas por endolinfa, existe una gran cantidad de K+ Parte inferior,
está bañada por perilinfa. Células ciliadas tienen un potencial intracelular de
-70 mV con respecto a la perilinfa, y de -150 mV respecto a la endolinfa.
Determinación de la frecuencia del sonido
Principio de
la posición→ Método
ampliado por el
SNC para detectar
las diferentes frecuencias
sonoras, determinando el punto de la
membrana basilar que se
estimula al máximo.
Principio
de la salva o de la frecuencia → Método para discriminar la frecuencia
sonoras bajas en el
intervalo de 20 hasta 2000
ciclos por segundo,
pueden provocar impulsos nerviosos sincronizados a la misma
frecuencia. Los impulsos viajan por el nervio
coclear hacia los núcleos cocleares y estos núcleos distinguen las
diversas frecuencias.
Determinación del volumen Sistema auditivo determina el
volumen de 3 formas:
Conforme el sonido se hace más fuerte, aumenta
la amplitud de vibración de membrana basilar y células ciliadas, por lo que se
excita las terminaciones nerviosas con más rapidez.
A medida que la amplitud aumenta, se estimula
más células ciliares a los márgenes de la porción resonante de la membrana
basilar, lo que produce sumación espacial de impulsos
Células
ciliadas externas se
estimulan considerablemente hasta
que la vibración
de membrana basilar alcanza gran intensidad
Ley de la potencia
Oído puede
discriminar diferencias en
la intensidad del
sonido como una
variación de aproximadamente
10000 veces. Así pues, la escala de intensidad está enormemente comprimida. Lo
que le permite a una persona interpretar diferencias en intensidades a través
de un intervalo mucho más
extenso que el que sería posible si no fuera por la
compresión en la escala de las intensidades.
Determinación de dirección del sonido
Una persona puede determinar la
dirección del sonido mediante 2 mecanismos principales:
Mediante el lapso transcurrido entre la entrada
del sonido en un oído y su entrada al lado opuesto (Este mecanismo es más
exacto).
Mediante las diferencias de intensidades del
sonido en los 2 oídos.
Mecanismos nerviosos
para detectar dirección
del sonido comienzan
en núcleos olivares superiores, el medial por lapso
entre señales que entran en los 2 oídos,
mientras que los laterales por diferencia de intensidad del sonido que llegan a
los 2 oídos.
Alteraciones de la audición
Sordera
Se lo divide en 2 tipos:
Sordera nerviosa→ Causada por la alteración de la cóclea o
circuitos del SNC del oído
Sordera de conducción→ Causada por afección de las estructuras
acústicas que transmiten el sonido hasta la cóclea.